Original Article - DOI:10.33594/000000605
Accepted 01 January 2023 - Published online 08 February 2023
Expression of the P53 Protein and Morphological Changes in Neurons in the Pyramidal Layer of the Hippocampus After Simulation of Surgical Interventions in the Nasal Cavity in Rats
bPeoples’ Friendship University of Russia, department of otorhinolaryngology, Moscow, Russia,
cPeoples’ Friendship University of Russia, Department of Disaster Medicine, Moscow, Russia,
dPeoples’ Friendship University of Russia, department of histology, cytology and embryology, Moscow, Russia,
eI.M. Sechenov First Moscow State Medical University (Sechenov University)¸ department of pathophysiology, Moscow, Russia
Keywords
Abstract
Background/Aims:
to determine the role of surgical stress on the formation of p53-positive and dark neurons (DN) in the hippocampus, and to examine the parallelism of their formation in the pyramidal layer of the hippocampus.Methods:
Simulated septoplasty was performed on 20 Wistar rats. The hippocampus and dentate gyrus (DG) were examined, in which the number of DN and p53-positive neurons was determined at 2, 4 and 6 days after surgery. In each rat, 10 brain slices were stained with antibodies to p53 protein with Meyer’s haematoxylin and 10 slices were stained with Nissl toluidine blue. Hippocampal subfields CA1, CA2, CA3 and DG were studied. In the pyramidal subfield layer, the absolute number of neurons that were nuclear antibody-positive to p53 protein was counted, as well as the number of dark neurons. The counting area in each subfield was 20934±1260 µm2. Neurons are counted using the Aperio ImageScope program. For the histological specimen analysis, the ImageJ software was used. The data obtained using cell counting methods were presented as mean ± SE. Then, they were compared between both groups using a t-test SPSS 21software.Results:
Compared with the control group (n = 5), the number of DN and p53-positive neurons increased in experimental animals at all periods. A direct relationship was obtained between the increase in the number of DN and p53-positive neurons in the hippocampal formation. Septoplasty simulation in rats results in the pathogenetic cascades onset, which, in its turn, changes the morpho-functional properties of neurons of the pyramidal layer of the hippocampus and contributes to their neuroplasticity. Activation of NMDS receptors of neurons during stress apparently, initiates two ways of neuron life – the beginning of p53 protein expression and the DN formation. Both ways can finally lead to apoptosis.Conclusion:
The formation of dark neurons and the expression of the p53 protein in them are most likely to be interconnected and can probably provide neuroprotective mechanisms.Introduction
Simulated septoplasty in rats leads to the development of a powerful stress response. Few experimental data are available to determine the consequences of nasal surgery [1].
Various stressors lead to changes in the functional state of neurons with the development of subsequent morpho-physiological abnormalities [2]. The hippocampus receives special attention in stress because it is very sensitive to various damaging factors [3, 4]. In neuronal damage, as in damage to other cells, p53 protein is an activator of transcription of a specific set of target genes, a cell cycle inhibitory regulatory factor and an effector of cellular responses to damage, which include cell cycle arrest and apoptosis [5]. However, p53 has also been shown to be neuroprotective. For example, this has been shown in an in vivo model of tautopathy [6]. P53 controls the transcription of a group of genes involved in the synaptic function of neurons. The transcriptional control of p53 of these synaptic genes is conserved in mouse neurons and the human brain [6]. Morphologically altered neurons after exposure to stressors can have basophilia in their staining [3, 7]. Such neurons are usually referred to as dark neurons. They have specific morphological features: shrunken cytoplasm, karyopiknosis, corkscrew axon [3, 7]. Apoptosis is believed to occur in these neurons [8]. However, it is not excluded that dark neurons are capable of restoring their morpho-functional state under certain conditions [7].
However, no studies evaluating the parallelism of p53 protein expression in hippocampal neurons and the appearance of dark neurons there in have been performed in a septoplasty simulation in rats.
The aim of this study was to determine the role of surgical stress on the formation of p53-positive and dark neurons in the hippocampus, and to examine the parallelism of their formation in the pyramidal layer of the hippocampus.
Materials and Methods
In the study, 20 sexually mature male Wistar rats weighing 250±20 g were randomly divided into experimental (n=15) and control (n=5) groups. The rats were kept under controlled temperature (23±2.5°C), 12-hour illumination and free access to water and food. Anesthesia was administered with zoletil 100 solution (15 mg/kg) 10 min before surgery to 15 rats, which constituted the experimental group.
The animals were housed in a specially equipped room, access to which was limited. Animals were housed in cages for individual housing. In the control group, there were rats that were anesthetized, as in experimental rats, but the simulation of septoplasty itself was not performed. Naive animals in the control group were excluded, since pharmacological substances for general anesthesia can change the morpho-functional state of neurons. All animals were housed in a separate room two weeks prior to surgery. In the experimental male rats were used no earlier than 2 weeks later – the period of adaptation to new conditions of detention. The rats received a standard diet once a day, with free access to water. All animals during the experiments were under the same conditions. In the postoperative period, all animals were constantly provided with food and clean water, as well as standard cage care. Additional methods of postoperative wound treatment were not used. The keeping of rats, modeling of surgical trauma – septoplasty, as well as the removal of animals from experience were carried out in accordance with the ethical standards set in the Geneva Convention “International Guiding principles for Biomedical Research Involving Animals” (Geneva, 1990).
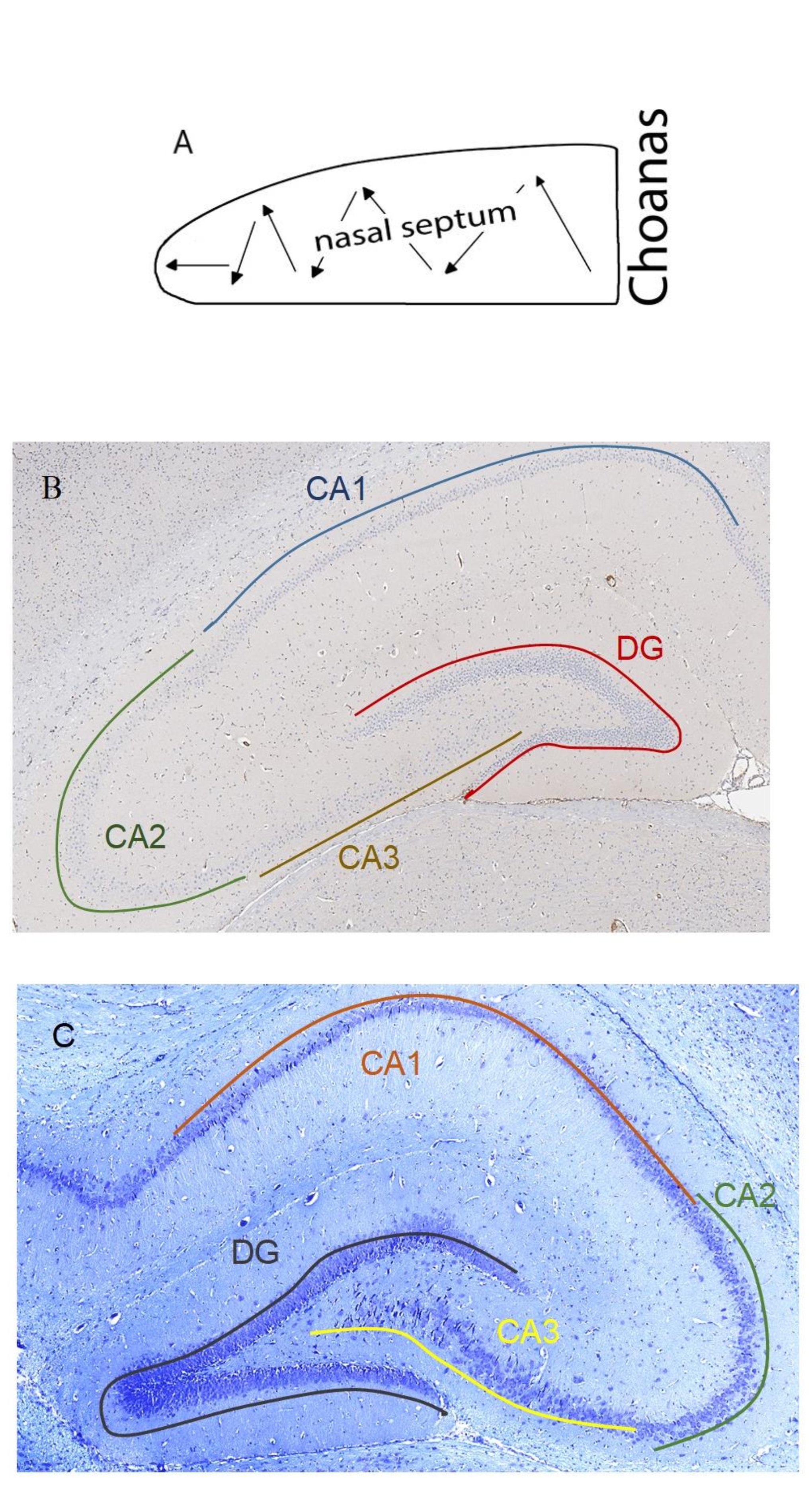
Fig. 1: А. Scheme of the septoplasty simulation. Arrows indicate the direction of the nasal septum scarification. B. Location of the rat hippocampus subfields. Immunohistochemical reaction anti-p53. Staining with Mayer's hematoxylin. Magnification, х10. С. Location of the rat hippocampus subfields. Nissl staining. Magnification,
Experimental procedure
20 animals in the experimental group were simulated septoplasty using the standard method by zigzag scarification of the nasal cavity mucosa with a sharp probe in caudo-cranial direction (Fig. 1a) [9]
Tissue probe
In the experimental and control groups, euthanasia was carried out by administering lethal doses of zoletil 100 solution. In the experimental group the animals were euthanised on the 2nd, 4th and 6th days postoperatively, 5 animals for each period. The brain was first perfused through the aorta with 0.9% sodium chloride solution, then with 10% formalin buffered solution, after which the brains were encased in paraffin blocks. Serial slices of the brain at the level of the bregma were taken with a microtome blade and 8 slices in the frontal plane, 4 µm thick, were obtained from each animal.
Staining
In each rat, 10 brain slices were stained with antibodies to p53 protein with Meyer’s haematoxylin and 10 slices were stained with Nissl toluidine blue. Hippocampal subfields CA1, CA2, CA3 and dentate gyrus (DG) were studied (Fig. 1b, 1с).
Morphometry
In the pyramidal subfield layer, the absolute number of neurons that were nuclear antibody-positive to p53 protein was counted, as well as the number of dark neurons (Fig. 2).
The counting area in each subfield was 20934±1260 µm2. Neurons are counted using the Aperio ImageScope program. For the histological specimen analysis, the ImageJ software was used.
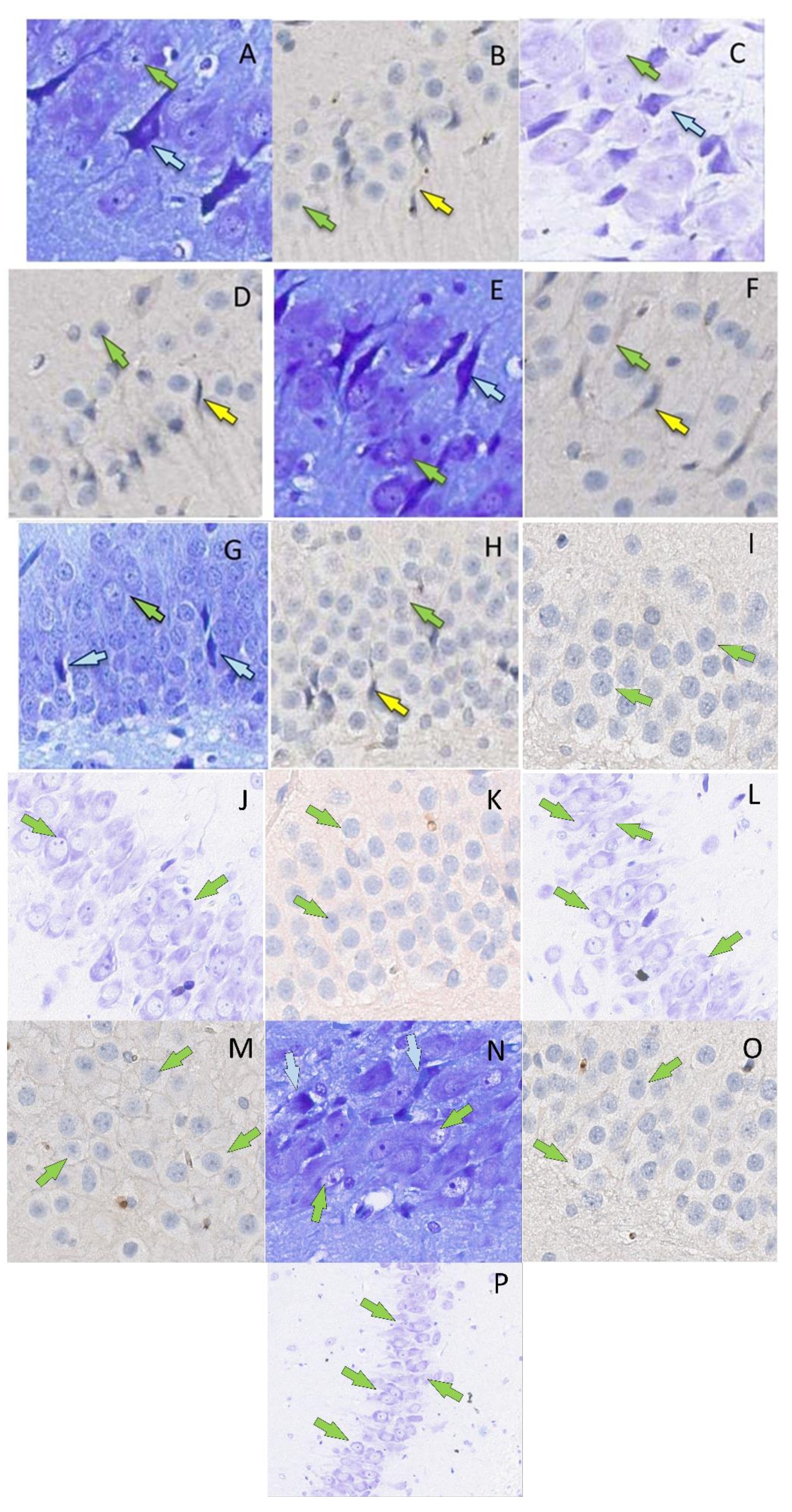
Fig. 2: P53-positive neurons (B, D, F, H, I, K, M, O) (yellow arrows, surrounded by mouse monoclonal antibodies to p53 protein, x400) and dark neurons (A, C, D, G, J, L, N, P) (blue arrows, Nissl toluidine blue, x400) in the hippocampal formation in rats on the 2nd (A, B, G), 4th (C, D, H) and 6th days (E, F) after the septoplasty simulation. Green arrows indicate intact neurons. Subfield CA1 – A, D, I, J; subfield CA2 – B, C, K, L; subfield CA3 – E, F, M, N; DG – G, H, O, P. Control group – I, J, K, L, M, N, O, P.
Statistical analysis of the data
The data obtained using cell counting methods were presented as mean ± SE. Then, they were compared between both groups using a t-test SPSS 21software.
Results
Number of dark neurons
There was a non-Gaussian distribution in the number of THs in the pyramidal layer of the hippocampus both in the experimental and control groups. In CA1, the number of DNs on days 2 and 4 did not differ significantly from the control, but on day 6 postoperatively there was a decrease (p<0.001) (Fig. 3b).
There was no significant difference in CA2 between the experimental group and the control group. On day 2 postoperatively, there was a minimum of THs compared to day 4 (p<0.001). On day 4 postoperatively, there was the peak in number of THs in CA3 compared to the rest of the day (p<0.001). In the control group, the number of THs was not different from day 2, but it was significantly lower compared to day 4 (p<0.001) and day 6 (p<0.05) after surgery (Fig. 3b). In DG, similar results to CA3 were observed (Fig. 3b) (Table 1).
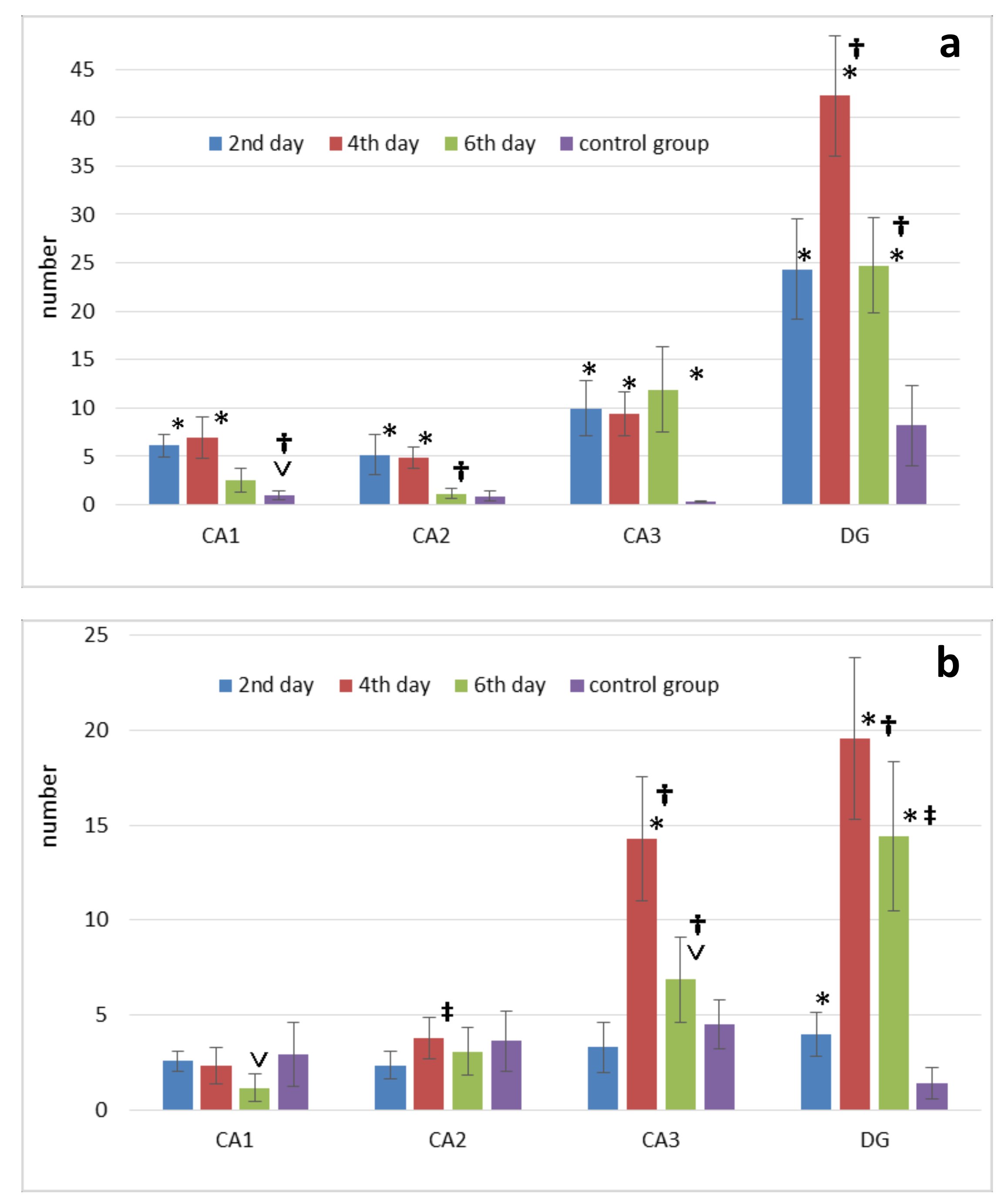
Fig. 3: Changes in the number of p53-positive neurons (p53) (a) and dark neurons (b) in the septoplasty simulation. Note: * - significant differences between data of the control group and terms after operation (p<0.001); ˅ - significant differences between data of the control group and terms after operation (p<0.05); † - significant differences between terms after operation within experimental group (p<0.001); ‡ - significant differences between terms after operation within the experimental group (p<0.05).
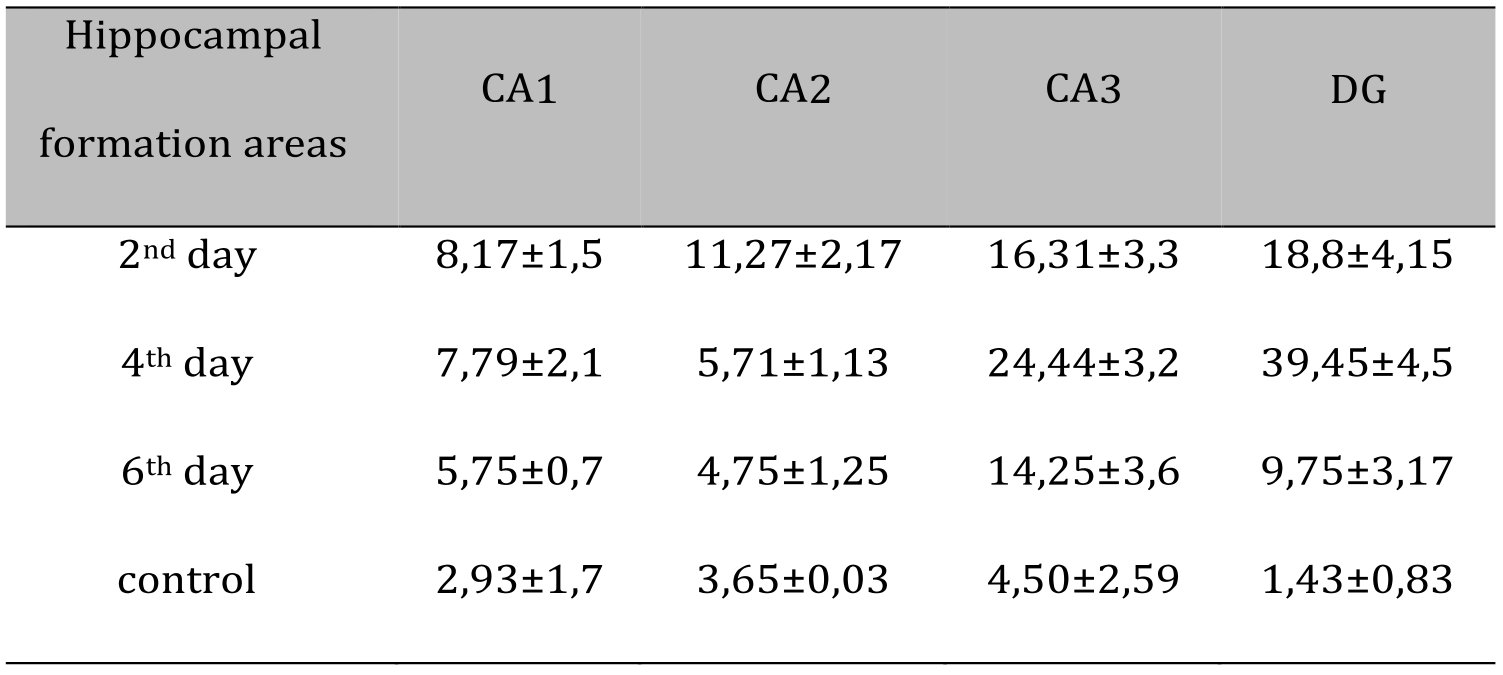
Table 1: DN number at different periods after septoplasty simulation in rats hippocampal formation by staining sections with Nissl toluidine blue stain
Number of p53-positive neuronss
ccording to Mann-Whitney test the number of p53-positive neurons in CA1 significantly increased on days 2, 4 (p<0.001) and 6 (p<0.05) after septoplasty compared to control group. Dynamically, the peak of increase in the p53 protein expression in the cytoplasm of CA1 and CA2 neurons of hippocampus occurred on days 2-4, and on day 6 the number of these neurons significantly decreased (p<0.001). On day 6, p53-positive neurons in CA2 did not differ from the control group (Fig. 3a). In CA3 there was an increase in p53 protein expression at all time points after surgery compared to the controls (p<0.001).
In DG, compared to the controls, the number of p53-positive neurons was significantly higher at all time points of evaluation. The number of these cells peaked on day 4, compared to the other terms (p<0.001) (Fig. 3a) (Table 2).
Comparing the number of neurons in which the p53 protein was expressed into the cytoplasm and the DN number, a positive strong correlation was found at all evaluation lines and in all hippocampal subfields (Fig. 4).
The lowest coefficient of determination was found when evaluating the CA2 subfield on the 4th day after surgery (Table 3).
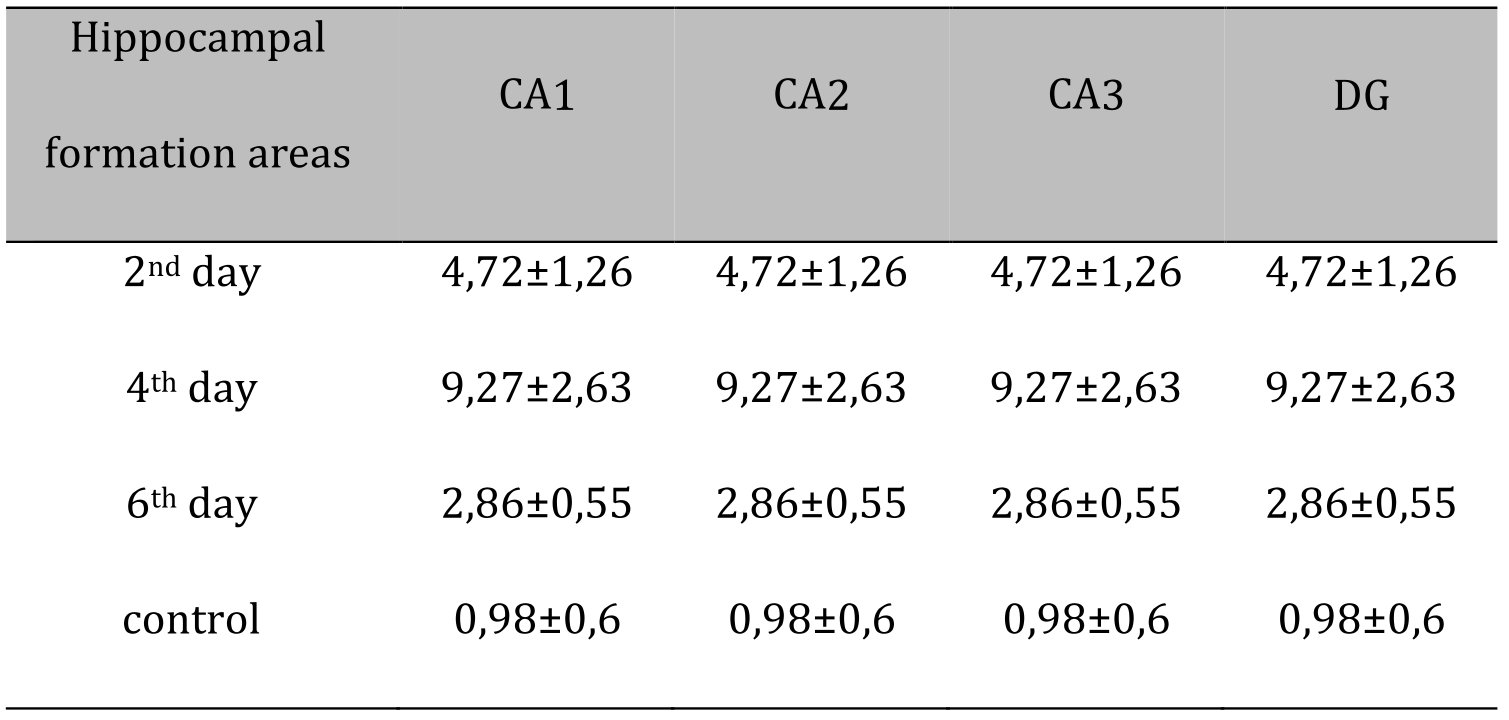
Table 2: Correlation coefficients between the concentrations of VDBP, AAT, and neutrophil granule proteins. Significant correlations (p<0.05) in bold type
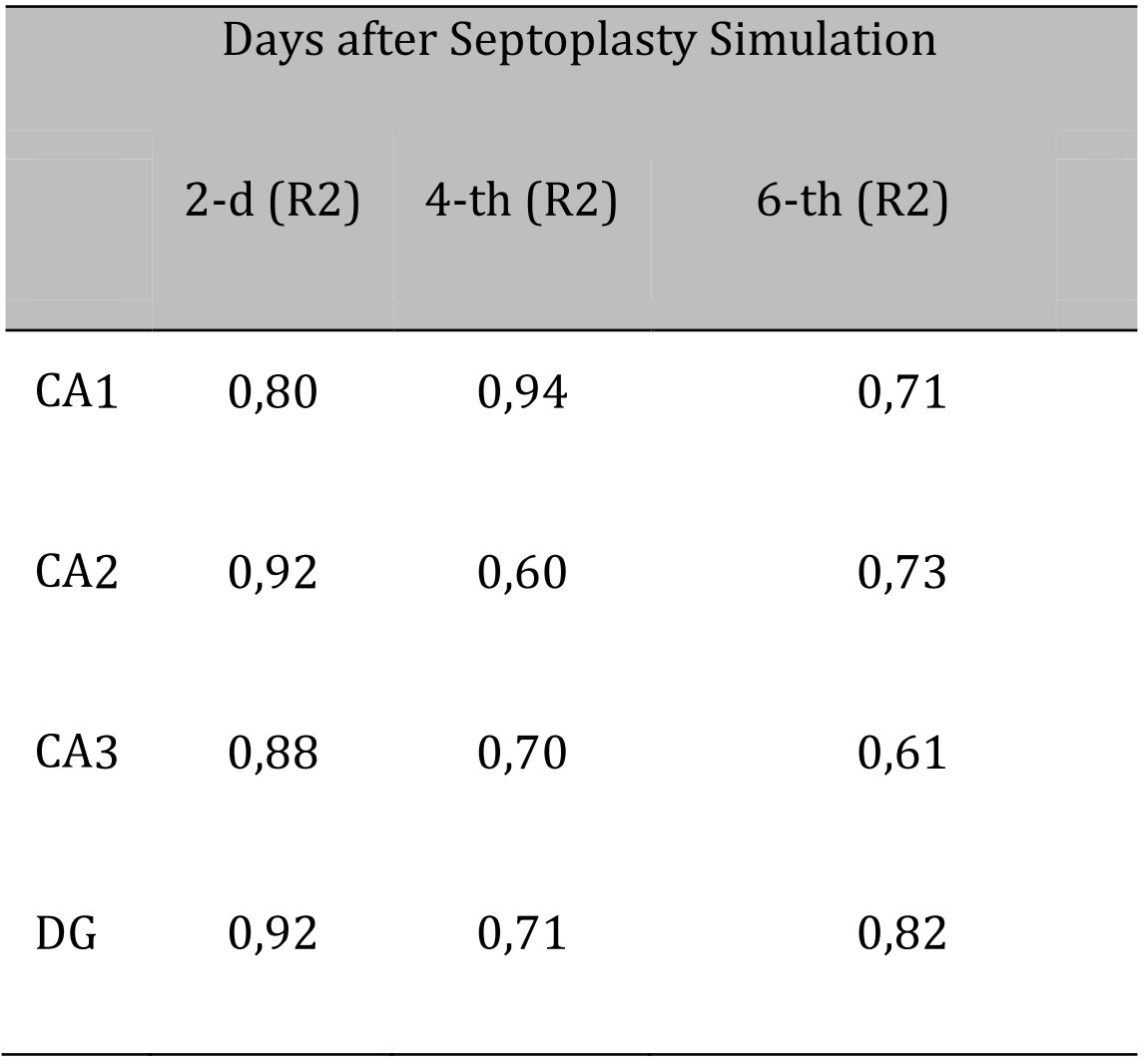
Table 3: Determination coefficients for comparing the number of dark neurons (DN) and the number of p53-positive neurons (p53) in the hippocampus after septoplasty simulation
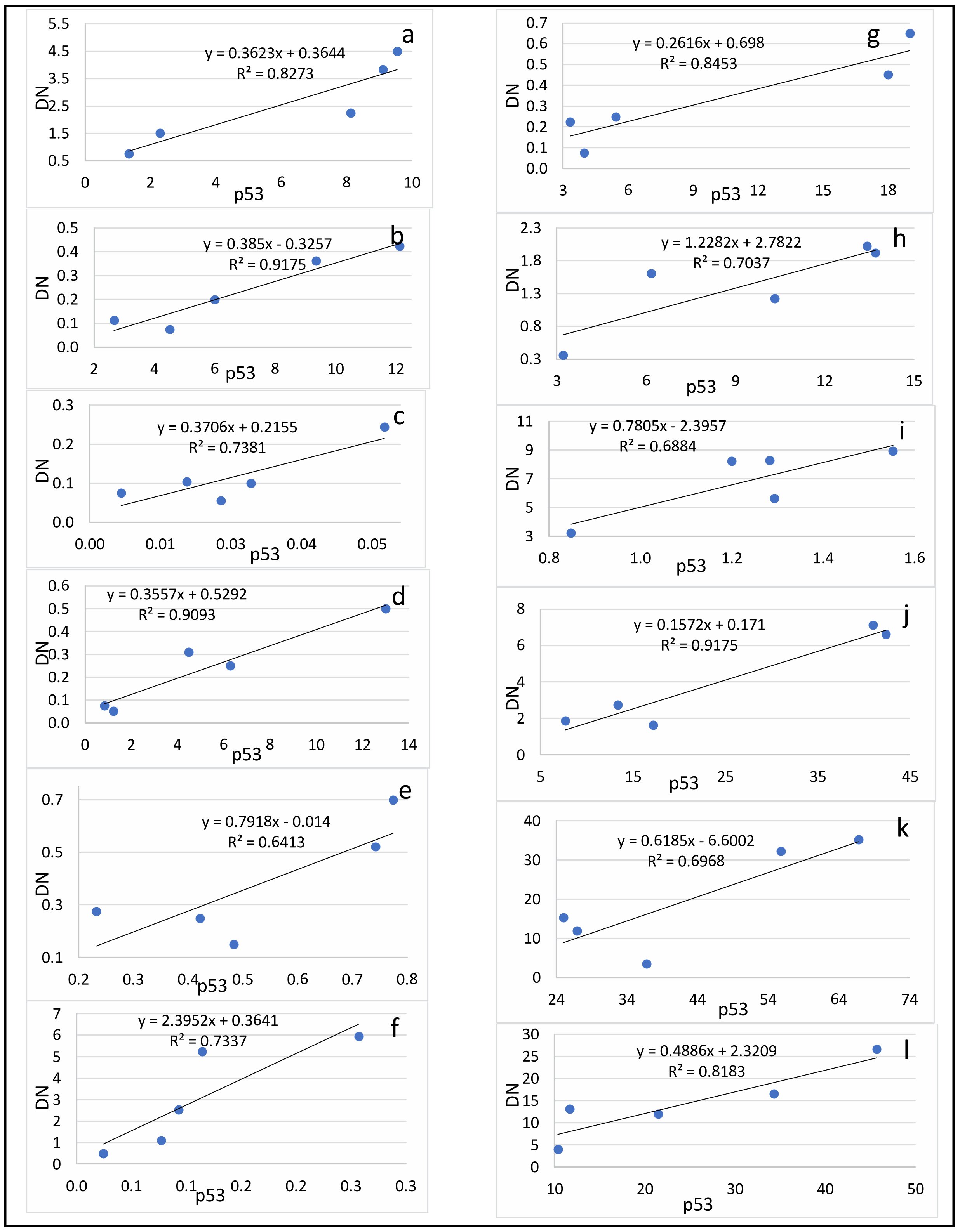
Fig. 4: Correlation between the number of dark neurons (DN) and the number of p53-positive neurons (p53) in the hippocampal subfields CA1 (a-c), CA2 (d-f), CA3 (g-i) and DG (j-l) on the 2nd (a, d, g, j), 4th (b, e, h, k) and 6th (c, f, i, l) days after the septoplasty simulation.
Discussion
The p53 protein is activated by cellular stress and DNA damage and, depending on the severity of stress and the specific cell type, can contribute to adaptive responses to stress or can trigger cell cycle arrest or its apoptosis [10]. When normal proliferating cells are DNA damaged, they may react in one of the two ways: cell cycle arrest r apoptosis, and p53 is engaged in both of those processes [11].
The p53 protein is an important component in the neuron apoptosis, for example, after ischemic event or excitotoxicity [12]. An increase in number of such neurons has been demonstrated in ischemia, traumatic brain injuries [13]. Some research studies have proved that p53 is part of the biochemical processes in the cell caused by the activation of NMDA receptors (N-methyl-D-aspartate) and finally resulting to apoptosis [14]
The mechanisms of proapoptotic action are assumed to be realized through the induction of the P53 gene (protein p53) expression, the regulation of which inhibits the passage of the cell cycle from G1 to S-phase, which blocks the division of cancer cells and tumor growth [15]. Except to the widely studied role of p53 as a regulator of apoptosis triggering, its neuroprotective role has also been demonstrated [16]. The main goal of neuroprotection is to prevent the death of neurons in the ischemic area, where apoptosis is one of the mechanisms of neuronal death. Bioenergetic processes are slowed down in the penumbra and neurons which have not died yet, remaining in them. In this regard, the absence of neurons in the hippocampus with obvious morphological signs of apoptosis in the analysis of the slices obtained by us, may indicate the presence of neuroprotective properties of the p53 protein. It has been shown that posttranslational modifications of p53 can contribute to the differentiation of neurons, as well as to the growth and regeneration of axons [17].
It was shown that p53 is a neuroprotector in an in vivo model of taupathy [18]. By analyzing the chromatin immunoprecipitation chip, it was determined, that p53 controls the transcription of a group of genes participated in providing synaptic function. Genetic manipulation of these genes changed the neurotoxicity of the tau- protein. The authors have found, that both in mice’s neurons and in the human brain, transcriptional control of these synaptic genes is maintained due to p53. Thus, it has been suggested, that the provision of synaptic function, as a manifestation of neuroprotection, can be performed by p53 protein [18].
In addition to activating the P53 gene, NMDA receptors participate in caspase-dependent apoptosis, increasing the level of calcium ions, boosting the enzyme caspase-3 activity. This enzyme, in turn, start the formation of dark neurons and their next degeneration. When inhibiting caspases with the pancaspase inhibitor FK011, a decrease in the changes characteristic of dark neurons has been achieved [19].
Previously, it was also shown, that dark neurons can both restore their morpho-functional state by increasing the cisterns of the granular endoplasmic reticulum with the formation of membrane curls, the transition of this process to astrocytic processes and, as a consequence, with a subsequent decline in the degree of structural compaction of the cell [7], and be a sign of the final necrotic decay of the cell regardless of the cause of neuron death, including various biochemical cascades of apoptosis [8]. There is an opinion, that dark neurons- are the result of oxidative stress. Thus, it has been shown, that the use of luteolin after brain injury in vivo reduces the number of dark neurons and oxidative stress in them in the hippocampus [20]. It has been shown, that the presence of regenerating dark neurons in the case of animal studies indicates the vulnerability of neuroprotective properties of neuroglia [21]. The presence of such phenomena, as cytoplasmic shrinkage and surface reduction in DN is compared by some authors with the manifestation of neuroplasticity characteristics [22, 23]. In fact, plasticity refers to the unique ability of the brain (neuron) to change and reorganize in response to changes in the environment. This property of neurons contributes to their viability and, consequently, the organism’s survival [24]. The most well-known examples of neuronal plasticity are the formation of new synapses, the proliferation of dendritic spines, the retraction and simplification of dendrites, and the reduction of dendritic spines under stressful conditions. Some studies have shown that endogenous or exogenous stressors are associated with a decrease in the surface and dendritic spike of neurons [25].
The high determination coefficients found in this study confirm the theory that presence of dark neurons in the hippocampus and dentate gyrus is most likely closely related to the expression of the p53 protein during surgical stress caused by septoplasty simulation in rats. This is probably due to the activation of NMDA receptors in neurons under the influence of surgical stress, as it has been shown that stress leads to an increase in the content of NMDA receptors in the dendritic spike apparatus [26]. In addition, exposure to a large amount of glutamate leads to functional changes in neurons and subsequent launch of the apoptosis program [27]. Stress is known to result in degeneration of hippocampal neuron dendrites [28]. In dark neurons, the dendrites are poorly developed or practically absent as a result of modulation of NMDA receptors. This was shown by the example of CA3 subfield neurons in the hippocampus [29]. In addition, chronic stress has been reported to cause atrophy of the pyramidal layer of the subfield CA1 [30, 31], decrease the long-term potentiation of neurons of the hippocampal CA1 subfield [32] and cause apoptosis of neurons, as well as a decrease in the density of dendritic spikes in neurons in the CA1 region of the hippocampus [33]. Thus, it can be assumed that there is a common mechanism that result to the start of two processes, discussed in this article – the expression of the p53 protein in the cytoplasm and the formation of dark neurons. The trigger of these pathways is probably the activation of NMDA receptors of neurons.
Morphological changes in the hippocamp are confirmed by our previous studies, which showed that septoplasty simulation in rats provokes the development of many stress reactions [34] and even a breakdown in adaptation [35]: an increase in the number of dark neurons [36], the expression of the p53 protein [3], a significant release of corticosterone into the blood plasma [37], and an increase in degranulation of mast cells [38], disturbances in the balance of the autonomic nervous system [37, 39, 40], changes in behavior when testing rats in an open field [9], the appearance of anxiety and a depressive-like state [37, 41-43].
Conclusion
Septoplasty simulation in rat’s results in the pathogenetic cascades onset, which, in its turn, changes the morpho-functional properties of neurons of the pyramidal layer of the hippocampus and contributes to their neuroplasticity. Activation of NMDA receptors of neurons during stress apparently, initiates two ways of neuron life - the beginning of p53 protein expression and the formation of dark neurons. Both ways can finally lead to apoptosis.
In further studies, it is necessary to study at what postoperative period the p53 protein expression in neurons decreases and the number of dark neurons in the hippocampal formation returns to normal values. Most likely, this should occur after the 14th day after surgery, since it is known that migrating neurons from the dentate gyrus to the hippocampus begin to synthesize neurotransmitters and fully perform their function starting from the 14th day of their appearance in the pyramidal layer of the hippocampus [19, 20, 32].
The formation of dark neurons and the expression of the p53 protein in them are most likely to be interconnected and can probably provide neuroprotective mechanisms.
Compliance with Ethical Standards
The keeping of rats, modeling of surgical trauma – septoplasty, as well as the removal of animals from experience were carried out in accordance with the ethical standards set in the Geneva Convention “International Guiding principles for Biomedical Research Involving Animals” (Geneva, 1990).
Disclosure Statement
The authors declare no conflicts of interest
References
| 1 | Dragunova SG, Reshetov IV, Kosyreva TF, Severin AE, Khamidulin GV, Shmaevsky PE, Inozemtsev AN, Popadyuk VI, Kastyro IV, Yudin DK, Yunusov TY, Kleyman VK, Bagdasaryan VV, Alieva SI, Chudov RV, Kuznetsov ND, Pinigina IV, Skopich AA, Kostyaeva MG. Comparison of the Effects of Septoplasty and Sinus Lifting Simulation in Rats on Changes in Heart Rate Variability. Dokl Biochem Biophys 2021;498:165-169. https://doi.org/10.1134/S1607672921030029 |
| 2 | Haider S, Naqvi F, Batool Z, Tabassum S, Perveen T, Saleem S, Haleem DJ. Decreased Hippocampal 5-HT and DA Levels Following Sub-Chronic Exposure to Noise Stress: Impairment in both Spatial and Recognition Memory in Male Rats. Sci Pharm 2012;80:1001. https://doi.org/10.3797/scipharm.1207-15 |
| 3 | Kastyro IV, Reshetov IV, Khamidulin GV, Shilin SS, Torshin VI, Kostyaeva MG, Popadyuk VI, Yunusov TY, Shmaevsky PE, Shalamov KP, Kupryakova AD, Doroginskaya ES, Sedelnikova AD. Influence of Surgical Trauma in the Nasal Cavity on the Expression of p53 Protein in the Hippocampus of Rats. Dokl Biochem Biophys 2021;497:99-103 https://doi.org/10.1134/S160767292102006X |
| 4 | Kirichuk VF, Tsymbal AA. Use of terahertz electromagnetic radiation at nitric oxide frequencies for the correction of thyroid functional state during stress. Vestn Ross Akad Med Nauk Russian 2010;4:37-40. |
| 5 | Sheahan S, Bellamy CO, Treanor L, Harrison DJ, Prost S. Additive effect of p53, p21 and Rb deletion in triple knockout primary hepatocytes. Oncogene 2003;23: 1489-1497. https://doi.org/10.1038/sj.onc.1207280 |
| 6 | Merlo P, Frost B, Peng S, Yang YJ, Park PJ, Feany M. p53 prevents neurodegeneration by regulating synaptic genes. Proc Natl Acad Sci U S A 2014;111:18055-18060. https://doi.org/10.1073/pnas.1419083111 |
| 7 | Csordás A, Mázló M, Gallyas F. Recovery versus death of "dark" (compacted) neurons in non-impaired parenchymal environment: light and electron microscopic observations. Acta Neuropathol 2003;106: 37-49. https://doi.org/10.1007/s00401-003-0694-1 |
| 8 | Kövesdi E, Pál J, Gallyas F. The fate of "dark" neurons produced by transient focal cerebral ischemia in a non-necrotic and non-excitotoxic environment: neurobiological aspects. Brain Res 2007;1147:272-283. https://doi.org/10.1016/j.brainres.2007.02.011 |
| 9 | Kastyro IV, Reshetov IV, Khamidulin GV, Shmaevsky PE, Karpukhina OV, Inozemtsev AN, Torshin VI, Ermakova NV, Popadyuk VI. The Effect of Surgical Trauma in the Nasal Cavity on the Behavior in the Open Field and the Autonomic Nervous System of Rats. Dokl Biochem Biophys 2020;492:121-123. https://doi.org/10.1134/S1607672920030023 |
| 10 | Jõers A, Jaks V, Kase J, Maimets T. p53-dependent transcription can exhibit both on/off and graded response after genotoxic stress. Oncogene 2004;23:6175-6185. https://doi.org/10.1038/sj.onc.1207864 |
| 11 | Bellamy CO. p53 and apoptosis. Br Med Bull 1997;53:522-538. https://doi.org/10.1093/oxfordjournals.bmb.a011628 |
| 12 | D'Orazi G. Recent Advances in p53. Biomolecules 2021;11:211. https://doi.org/10.3390/biom11020211 |
| 13 | Filichia E, Shen H, Zhou X, Qi X, Jin K, Greig N, Hoffer B, Luo Y. Forebrain neuronal specific ablation of p53 gene provides protection in a cortical ischemic stroke model. Neuroscience 2015;295:1-10. https://doi.org/10.1016/j.neuroscience.2015.03.018 |
| 14 | Miller FD, Pozniak CD, Walsh GS. Neuronal life and death: an essential role for the p53 family. Cell Death Differ 2000;7:880-888. https://doi.org/10.1038/sj.cdd.4400736 |
| 15 | Rivlin N, Brosh R, Oren M, Rotter V. Mutations in the p53 Tumor Suppressor Gene: Important Milestones at the Various Steps of Tumorigenesis. Genes Cancer 2011;2:466-474. https://doi.org/10.1177/1947601911408889 |
| 16 | Khurana V, Merlo P, DuBoff B, Fulga TA, Sharp KA, Campbell SD, Götz J, Feany MB. A neuroprotective role for the DNA damage checkpoint in tauopathy. Aging Cell 2012;11:360-362. https://doi.org/10.1111/j.1474-9726.2011.00778.x |
| 17 | Tedeschi A, Di Giovanni S. The non-apoptotic role of p53 in neuronal biology: enlightening the dark side of the moon. EMBO Rep 2009;10:576-583. https://doi.org/10.1038/embor.2009.89 |
| 18 | Merlo P, Frost B, Peng S, Yang YJ, Park PJ, Feany M. p53 prevents neurodegeneration by regulating synaptic genes. Proc Natl Acad Sci U S A 2014;111:18055-18060. https://doi.org/10.1073/pnas.1419083111 |
| 19 | Coles J, Oomman SK, Henne WM, Bliss RM, Hoffman TC, Finckbone V, Strahlendorf H, Strahlendorf J. AMPA-induced dark cell degeneration is associated with activation of caspases in pyramidal neurons of the rat hippocampus. Neurosci Lett 2008;436:294-299. https://doi.org/10.1016/j.neulet.2008.02.075 |
| 20 | Ashaari Z, Hassanzadeh G, Mokhtari T, Hosseini M, Keshavarzi Z, Amini M, et.al. Luteolin Reduced the Traumatic Brain Injury-Induced Memory Impairments in Rats: Attenuating Oxidative Stress and Dark Neurons of Hippocampus. Acta medica Iranica 2018;56:563-570. |
| 21 | Gallyas F, Hsu M, Buzsáki G. Four modified silver methods for thick sections of formaldehyde-fixed mammalian central nervous tissue: 'dark' neurons, perikarya of all neurons, microglial cells and capillaries. J Neurosci Methods 1993;50:159-164. https://doi.org/10.1016/0165-0270(93)90004-B |
| 22 | Kherani ZS, Auer RN. Pharmacologic analysis of the mechanism of dark neuron production in cerebral cortex. Acta Neuropathol 2008;116:447-452. https://doi.org/10.1007/s00401-008-0386-y |
| 23 | Ahmadpour S, Behrad A, Fernández-Vega I. Dark Neurons: A protective mechanism or a mode of death. Journal of Medical Histology 2019;3:125-131. https://doi.org/10.21608/jmh.2020.40221.1081 |
| 24 | Marder E, Prinz AA. Current compensation in neuronal homeostasis. Neuron 2003;37:2-4. https://doi.org/10.1016/S0896-6273(02)01173-X |
| 25 | McEwen BS, Nasca C, Gray JD. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 2016;41:3-23. https://doi.org/10.1038/npp.2015.171 |
| 26 | Sun DS, Zhong G, Cao HX, Hu Y, Hong XY, Li T, Li X, Liu Q, Wang Q, Ke D, Liu GP, Ma RH, Luo DJ. Repeated Restraint Stress Led to Cognitive Dysfunction by NMDA Receptor-Mediated Hippocampal CA3 Dendritic Spine Impairments in Juvenile Sprague-Dawley Rats. Front Mol Neurosci 2020;13:552787. https://doi.org/10.3389/fnmol.2020.552787 |
| 27 | Elhussiny MEA, Carini G, Mingardi J, Tornese P, Sala N, Bono F, Fiorentini C, La Via L, Popoli M, Musazzi L, Barbon A. Modulation by chronic stress and ketamine of ionotropic AMPA/NMDA and metabotropic glutamate receptors in the rat hippocampus. Prog Neuropsychopharmacol Biol Psychiatry 2020;104:110033. https://doi.org/10.1016/j.pnpbp.2020.110033 |
| 28 | Xu Y, Pan J, Sun J, Ding L, Ruan L, Reed M, Yu X, Klabnik J, Lin D, Li J, Chen L, Zhang C, Zhang H, O'Donnell JM. Inhibition of phosphodiesterase 2 reverses impaired cognition and neuronal remodeling caused by chronic stress. Neurobiol Aging 2015;36:955-970. https://doi.org/10.1016/j.neurobiolaging.2014.08.028 |
| 29 | Christian KM, Miracle AD, Wellman CL, Nakazawa K. Chronic stress-induced hippocampal dendritic retraction requires CA3 NMDA receptors. Neuroscience 2011;174: 26-36. https://doi.org/10.1016/j.neuroscience.2010.11.033 |
| 30 | Sandi C, Pinelo-Nava MT. Stress and memory: behavioral effects and neurobiological mechanisms. Neural Plast 2007;2007:78970. https://doi.org/10.1155/2007/78970 |
| 31 | Lupien SJ, McEwen BS, Gunnar MR, Heim C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat Rev Neurosci 2009;10:434-445. https://doi.org/10.1038/nrn2639 |
| 32 | Bhagya VR, Srikumar BN, Veena J, Shankaranarayana Rao BS. Short-term exposure to enriched environment rescues chronic stress-induced impaired hippocampal synaptic plasticity, anxiety, and memory deficits. J Neurosci Res 2017;95:1602-1610. https://doi.org/10.1002/jnr.23992 |
| 33 | Huang P, Li C, Fu T, Zhao D, Yi Z, Lu Q, Guo L, Xu X. Flupirtine attenuates chronic restraint stress-induced cognitive deficits and hippocampal apoptosis in male mice. Behav Brain Res 2015;288:1-10. https://doi.org/10.1016/j.bbr.2015.04.004 |
| 34 | Kastyro IV, Popadyuk VI, Muradov GM, Reshetov IV. Low-Intensity Laser Therapy As a Method to Reduce Stress Responses after Septoplasty. Dokl Biochem Biophys 2021;500:300-303. https://doi.org/10.1134/S1607672921050112 |
| 35 | Kastyro IV, Reshetov IV, Popadyuk VI, Torshin VI, Ermakova NV, Karpukhina OV, Inozemtsev AN, Khamidulin GV, Shmaevsky PE, Sardarov GG, Gordeev DV, Scopich AA. Studying the physiological effects of a new model of septoplasty in rats. Head and Neck Russian Journal Russian 2020;8:33-38. |
| 36 | Kostyaevaa MG, Kastyro IV, Yunusov TYu, Kolomin TA, Torshin VI, Popadyuk VI, Dragunova SG, Shilin SS, Kleiman VK, Slominsky PA, Teplov AY . Protein p53 Expression and Dark Neurons in Rat Hippocampus after Experimental Septoplasty Simulation. Molecular Genetics, Mol Genet Microbiol 2022;37:19-24. https://doi.org/10.3103/S0891416822010037 |
| 37 | Kastyro IV, Popadyuk VI, Reshetov IV, Kostyaeva MG, Dragunova SG, Kosyreva TF, Khamidulin GV, Shmaevsky PE. Changes in the Time-Domain of Heart Rate Variability and Corticosterone after Surgical Trauma to the Nasal Septum in Rats. Dokl Biochem Biophys 2021;499:247-250. https://doi.org/10.1134/S1607672921040098 |
| 38 | Kastyro I., Kostyaeva M., Dragunova S., Kosyreva A. Effect of blood corticosterone concentration on mast cell degranulation in the mesentery in rats after maxillofacial surgical trauma. Virchows Archiv 2021;479. |
| 39 | Dragunova SG, Reshetov IV, Kosyreva TF, Severin AE, Khamidulin GV, Shmaevsky PE, Inozemtsev AN, Popadyuk VI, Kastyro IV, Yudin DK, Yunusov TY, Kleyman VK, Bagdasaryan VV, Alieva SI, Chudov RV, Kuznetsov ND, Pinigina IV, Skopich AA, Kostyaeva MG. Comparison of the Effects of Septoplasty and Sinus Lifting Simulation in Rats on Changes in Heart Rate Variability. Dokl Biochem Biophys 2021;498:165-169. https://doi.org/10.1134/S1607672921030029 |
| 40 | Dolgalev AA, Svyatoslavov DS, Pout VA, Reshetov IV, Kastyro IV. Effectiveness of the Sequential Use of Plastic and Titanium Implants for Experimental Replacement of the Mandibular Defect in Animals Using Preliminary Digital Design. Dokl Biochem Biophys 2021;496: 36-39. https://doi.org/10.1134/S160767292101004X |
| 41 | Kastyro IV, Shmaevsky PE, Khamidullin GV, Torshin VI, Kovalenko АN, Guseinov II, Pryanikov PD, Inozemtsev AN. The impact of trauma of the mucous membrane of the nasal septum in rats on behavioral responses and changes in the balance of the autonomic nervous system (pilot study). J Phys Conf Ser 2020;1611. https://doi.org/10.1088/1742-6596/1611/1/012054 |
| 42 | Torshin VI, Kastyro IV, Reshetov IV, Kostyaeva MG, Popadyuk VI. The Relationship between p53-Positive Neurons and Dark Neurons in the Hippocampus of Rats after Surgical Interventions on the Nasal Septum. Dokl Biochem Biophys 2022;502:30-35. https://doi.org/10.1134/S1607672922010094 |
| 43 | Dragunova SG, Kosyreva TF, Khamidulin GV, Shmaevsky PE, Ermakova NV, Severin AE, Torshin VI, Kastyro IV, Scopich AA, Gordeev DV, Sedelnikova AD, Kuznetsov ND, Popadyuk VI, Yudin DK. Assessment of the impact of closed sinus lifting on changes in the autonomic nervous system in the early postoperative period. Head and neck Russian Journal Russian 2022;10:8-15. |