Original Article - DOI:10.33594/000000731
Accepted 17 September 2024 - Published online 6 October 2024
Change in Nfkb/Nrf2/Bax Levels by High Monomeric Polyphenols Berries Extract (HMPBE) in Acute and Chronic Secondary Brain Damage
bDepartment of Veterinary Sciences, University of Messina, Viale SS Annunziata, 98168 Messina, Italy,
cDepartment of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Messina, Viale F. Stagno D’Alcontres 31, 98166 Italy,
dDepartment of Pharmaceutical Chemistry, College of Pharmacy, King Saud University, Riyadh 11451, King-dom of Saudi Arabia,
eInstitut für Ernährungswissenschaft, Justus Liebig Universitat Giessen, 35390 Giessen, Germany,
fHealth Care AG Zurich, Switzerland,
gNAM-Institute, Salzburg, Salzburg, A-5020, Austria,
hDepartment of Bio-science and Engineering, Faculty of System Science and Engineering, Shibaura Institute of Technology, Japan,
iDepartment of Biomedical, Dental and Morphological and Functional Imaging University of Messina, Via Consolare Valeria, 98125 Messina, Italy
Keywords
Abstract
Background/Aims:
High Monomeric Polyphenols Berries Extract (HMPBE) is a formula highly rich in polyphenols clinically proven to enhance learning and memory. It is currently used to enhances cognitive performance including accuracy, working memory and concentration.Methods:
Here, we investigated for the first time the beneficial effects of HMPBE in a mouse model of acute and chronic traumatic brain injury (TBI).Results:
HMPBE, at the dose of 15 mg/kg was able to reduce histological alteration as well as inflammation and lipid peroxidation. HMPBE ameliorate TBI by improving Nrf-2 pathway, reducing Nf-kb nuclear translocation and apoptosis, and ameliorating behavioral alteration such as anxiety and depression. Moreover, in the chronic model of TBI, HMPBE administration restored the decline of Tyrosine Hydroxylase (TH) and dopamine transporter (DAT) and the accumulation of a-synuclein into the midbrain region. This finding correlates the beneficial effect of HMPBE administration with the onset of parkinsonism related to traumatic brain damage.Conclusion:
The data may open a window for developing new support strategies to limit the neuroinflammation event of acute and chronic TBI.Introduction
Traumatic brain injury (TBI) is the term used to describe a variety of violent or sustained mechanical impacts that cause vascular injury and hypoxia, which in turn will cause inflammation, cell death, and tissue loss [1]. TBI can result in a variety of neurological problems, including disabilities and death, and it significantly burdens affected families and society. The World Health Organization (WHO) states that TBI is a serious issue that affects millions of people and is a major cause of mortality and disability in the US, where one in three injuries result in a TBI [2]. Three phases of TBI sequelae can be distinguished: acute (0–1 week), post–acute (1week–1 month), and chronic (1 month to years). Cell necrosis from direct force transfer to the brain tissue occurs during the acute phase, followed by secondary cell death from axonal disease and inflammation [3, 4]. Although secondary cell death and inflammation result from necrosis of the cells, post-acute and chronic phases shows signs of neuronal remodeling and chronic pathology, including neurodegeneration, ongoing inflammation, and protein misfolding of proteins like α-syn, APP, Tau, and transactive response DNA binding protein 43 kDa [3-7]. This point is probably the key of the substantial biochemical connection between the etiology of TBI and the onset of parkinsonism in humans. As a result, a great deal of current research attempts to fully explore this pathological overlap and take advantage of the shared targets between PD and TBI in order to create more potent and long-lasting treatment plans. The pathophysiology of Parkinson’s disease (PD) is commonly linked to the neuropathological mechanism of α-syn overexpression and Lewy body accumulation in SN [8]. The 14-kDa protein α-syn regulates synaptic activity through the trafficking control of vesicle docking and fusion as well as neurotransmitter release. α-syn immunohistochemistry is considered as one of the most reliable methods for assessing the neuropathology of Parkinson’s disease [9-11]. Moreover, an abnormal production of reactive oxygen species (ROS) by mitochondrial dysregulation and the altered expression of tyrosine hydroxylase (TH) positive dopaminergic neurons present in substantia nigra is modulated by the elevated upregulation of activated microglia are a connection point between TBI and PD onset. Stress signals sent by neurons upon injury activates astrocytes and microglia which consequently releases pro inflammatory factors such as tumor necrosis factor α (TNFα) and IL-1β [12]. This further recruits microglia to the injury site in order to clear cellular debris and favor the synthesis of neurotrophic factors. On the other hand, the increased upregulation of pro- inflammatory factors causes mitochondrial stress by production of ROS [6]. α-syn that has been misfolded and comes from injured neurons sets off a cascade of signaling pathways in glial and neuronal cells. While the nuclear factor KB (NF-ΚB) signaling pathway, many proinflammatory cytokines are produced and expressed when such events are activated. As a result, this series of events exacerbates neurodegenerative processes, especially in diseases like Parkinson’s disease and synucleinopathies. Among the numerous cells and tissues that express the NF-ΚB family of inducible transcription factors there are microglia, astrocytes, and neurons. Although the traditional NF-ΚB pathway is vital for controlling the synthesis of inflammatory mediators during neuroinflammation, the dual role of NF-KB activation can be critical for neuronal survival. Mitigating dopaminergic neuronal degeneration and Parkinson’s disease (PD) may be possible by selectively acting on NF-ΚB-associated pathways. By selectively altering the NF-ΚB-mediated signaling pathways, a number of naturally occurring chemicals found in medicinal plants can be a useful treatment option in attenuating dopaminergic neuronal loss associated with Parkinson’s disease [13]. Unfortunately, palliative care is the only realistic treatment available right now for persistent TBI and PD. Therefore, investigations targeted at finding substances capable of reducing or preventing PD in at-risk groups need to understand the molecular link between TBI and PD. More and more studies in recent years evaluated the beneficial effects of dietary supplement or nutrients on brain during inflammatory or oxidative stress condition [14-17]. Fruits and vegetables high in polyphenols are now extensively examined for their possible positive benefits as “functional foods” promoting a healthy brain and a fast recovery after damages [18].
Researchers have recently become interested in flavonoids due to their strong antioxidant qualities and exceptional anti-neuroinflammatory activity in a variety of neurodegenerative diseases, including Parkinson’s disease. Numerous flavonoid subclasses, such as flavones, flavonols, isoflavones, and anthocyanins, have been investigated for their potential to protect against both in vitro and in vivo Parkinson’s disease models. The evidence currently available is compelling for the idea that bioactive flavonoids could be used as lead pharmacophores or as an ingredient in food to treat Parkinson’s disease (PD) caused by inflammation [13].
Previous research has demonstrated that consuming blueberry or grape juice every day for a period of 12 weeks improved older volunteers’ episodic memory abilities [19]. More recently, Bensalem et al. demonstrated that, after 8 weeks of supplementation, the combination of a polyphenol-rich grape and blueberry extract was able to prevent age-related memory decline in middle-aged mice (16 months old) [18]. Additionally, Dudonnè et al, in recent bioavailability study found that blueberry phenolic metabolites increased in mouse plasma when co-ingested with polyphenol-rich grape extract emphasized the synergistic potential of grape and blueberry phenolic compounds. The same study also showed that, in comparison to a single acute dosage, long-term administration of the grape-blueberry combination considerably boosted plasma phenolic concentrations. With the addition of individual extracts, this result was not seen [20]. Furthermore, Impellizzeri et al. explored the effect of MemophenolTM, a compound rich in polyphenols derived from French grape (Vitis vinifera L.) and wild North American blueberry (Vaccinium angustifolium A.) extracts in an experimental model of AlCl3-induced Alzheimer Disease (AD). They focused on the oxidative stress and inflammatory processes demonstrated its ability to control symptoms of AD, including aberrant overexpression of APP, accumulation of β-amyloid, behavioral changes related to cognition and memory, and phosphorylated Tau levels [21].. Based on the polyphenol content listed below, this fraction of High Monomeric Polyphenols Berries Extract (HMPBE) contains most equal amounts of flavanol monomers and oligomers as similar to cocoa. Recent studies on large-scale flavanol derived from cocoa ingestion have indicated that the long-term consumption of flavanol fractions enhances cognitive functions that depend on the hippocampus [22, 23]. HMPBE is a special combination of extracts that offers a special ratio of carefully chosen polyphenols with synergistic effects on cognition that have been shown in studies to assist both short- and long-term memory and improve learning ability. HMPBE influences the two key factors that affect memory: neurogenesis and synaptic plasticity [22, 23]. HMPBE is rich in:
- Total flavonoids (flavan-3-ols, flavonols and anthocyanins): >43%
- Flavan-3-ols monomers: ≥20%
- Oligomers (DP ≤4): ≥22%
- Flavonols (quercetin, glycosylated derivatives): ≥ 0.15%
- Anthocyanins: ≥ 0.10%
Materials and Methods
Animals
CD1 male mice (8-week-old, 18-24g) were acquired from Envigo (Milan, Italy) and located in a controlled environment and provided with standard rodent chow (Teklad standard diet acquire from Envigo) and water available ad libitum. They were housed 5 mice/cage and maintained in a 12:12 h light–dark cycle at 21 ± 1°C and 50±5% humidity. The University of Messina Review Board for animal care (OPBA) approved the study.
Experimental design and groups
- First experimental design
To find the lowest dose with a significant effect we administered HMPBE dissolved in saline orally at three different increasing concentrations (5-15-30 Mg/kg) over a period of 3 months. At the end of the period, the animals were subjected to behavioural analysis as described below. At the end of the first experiment, 15 mg/kg was selected as the minimum dose to have an effect.
- Second experimental design
The controlled impactor device Impact OneTM Stereotaxic impactor for controlled cortical impact (CCI 9(Leica, Milan, Italy) was used to create a cortical contusion on the exposed cortex after a craniotomy (tip diameter: 4 mm; cortical contusion depth: 3 mm; impact velocity: 1.5 m/s). Sham mice underwent the identical surgical procedure but were not injured [24-26].
Mice were divided as following:
- Sham + vehicle group: mice were subjected to the surgical procedures as above except that the impact was not applied and animals were treated with oral somministration (o.s.) with vehicle (data not shown);
- Sham + HMPBE: mice were subjected to the surgical procedures as above except that the impact was not applied and animals were treated o.s. with HMPBE;
- TBI: mice were subjected to CCI plus administration of vehicle (saline);
- TBI + HMPBE: As for the TBI + vehicle group but HMPBE was administered o.s. at 15 mg/kg in saline for 1h after TBI;
- Third experimental design
Mice were subjected to TBI as previously described but they were sacrificed 30 days after surgical procedures [27].
Mice were divided as following:
- Sham + vehicle group: mice were subjected to the surgical procedures as above except that the impact was not applied and animals were treated o.s. with vehicle for 30 days (data not shown);
- Sham + HMPBE: mice were subjected to the surgical procedures as above except that the impact was not applied and animals were treated o.s. with HMPBE for 30 days;
- TBI: mice were subjected to CCI plus administration of vehicle (saline) for 30 days;
- TBI + HMPBE: As for the TBI + vehicle group but HMPBE was administered o.s. at 15 mg/kg in saline for 1h and once a day for 30 days after TBI;
Considering that we don’t find any significantly difference between Sham and Sham+HMPBE group we choose to show only Sham+ HMPBE.
Behavioural analysis
During the different experiment, designed group of animals underwent behavioural testing at 1 and 30 days. Mice were moved to the behaviour testing room 30 minutes before the first trial started so they could become accustomed to the environment. Based on behavioural tests that were used to keep the environment as uniform as feasible, animals were trained to use the equipment before every recording. The behavioural tests were conducted by three distinct trustworthy experts who were blinded to the animals’ damage state. Below a brief description of tests:
Histological brain analysis
After the experiment, brain tissue was removed, fixed at room temperature in buffered formaldehyde solution (10% in phosphate buffered saline), dehydrated by graduated ethanol, and then embedded in paraffin. Light microscopy was used to examine tissue sections that were 7 um thick after being deparaffinized with xylene and stained with haematoxylin/eosin (Bio-Optica, Milan, Italy). The number of damaged neurons was counted, and the gray matter’s histopathologic alterations were graded on a 6-point scale : No lesion was found, 1; 1–5 eosinophilic neurons were present in the gray matter, 2; 5–10 eosinophilic neurons were present, 3; more than 10 eosinophilic neurons were present, 4; a small infarction (less than one third of the gray matter area), 5; a moderate infarction (one third to one half of the gray matter area); and 6, a large infarction (more than half of the gray matter area). To determine a final score for each mouse, the results from every part of each brain were averaged. The slices were then analysed by a blinded histopathologist using an optical microscope using a Leica DM6 microscope (Leica Microsystems Spa, Milan, Italy)[42].
Western Blot Analysis of Cytosolic and Nuclear Extracts
Cytosolic and nuclear extracts were prepared as previously described [43]. The following primary antibodies were used: anti-Ikb-α (1:500; Santa Cruz Biotechnology (SCB), H-4 sc-1643), anti-Nf-kb p65 (1:500; SCB F-6: sc-8008), anti-Bax (1:500; SCB, B-9 sc-7480), anti-Bcl-2 (1:500; SCB, C-2 sc-7382), anti-a-syn (1:500SCB, LB509 sc-58480), anti-Nrf-2 (1:500; SCB, A-10 sc-365949), anti-HO-1 (1:500; SCB, A3 sc-136960), anti-DAT (1:300; SCB, 65G10 sc-32258) in 1× PBS, 5% w/v non-fat dried milk, 0.1% Tween-20 at 4 °C overnight. Blots were further probed with an anti—b-actin protein antibody (1:500; SCB) for the cytosolic fraction or an anti-lamin A/C antibody (1:500 Sigma-Aldrich Corp., Milan, Italy) for the nuclear fraction to make sure that they were loaded with an equivalent number of proteins [44, 45]. As directed by the manufacturer, signals were evaluated using an enhanced chemiluminescence (ECL) detection system reagent (Thermo, Monza, Italy). Using BIORAD ChemiDoc TM XRS+ software and densitometry, the relative expression of the protein bands was measured and standardised to the levels of b-actin and lamin A/C.
Cytokines measurement
Using commercially available enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems, Minneapolis, MN, USA) in accordance with the manufacturer’s instructions, TNF-a, IL-1b, and IL-6 levels from brain were measured as previously described [46].
Immunohistochemical localization of TH
The immunohistochemical techniques used have been previously described [47]. Briefly, slices were incubated with anti-TH (Millipore, 1:500 in PBS, v/v) for the entire night. After that, sections were treated with bovine anti-mouse IgG secondary antibodies (1:2, 000 Jackson Immuno Research, West Grove, PA, USA). Bovine anti-mouse immunoglobulin G (IgG) secondary antibody conjugated with peroxidase (1:2000 Jackson Immuno Research, West Grove, PA, USA) was applied to the sections after they had been cleaned with PBS. Using avidin-biotin peroxidase combination and goat anti-mouse IgG conjugated with biotin (Vector Laboratories, Burlingame, CA, USA), specific labelling was observed. Control slices were incubated with either primary or secondary antibody, none of which produced positive staining, to confirm the specificity of antibody-binding. The digital photos were first opened in ImageJ, then the colour deconvolution plug-in was used to deconvolute them. The deconvoluted DAB image is automatically plotted into a histogram profile when the IHC profiler plug-in is used, and a matching scoring log is shown. The positive pixel intensity value returned from the computer software matches the histogram profile. Two investigators who were blinded to the treatment performed all immunohistochemistry analyses [48, 49].
Malondialdehyde measurement
Hank’s balanced salt solution was homogenized at 3000 rpm with brain tissues present. For one hour at 95 degrees, pellets were incubated in a solution containing water, acetic acid, sodium dodecyl sulfate, and thiobarbituric acid. After cooling, the mixture was combined with water, n-butanol, and pyridine, then centrifuged at 3000 rpm. At 532 nm, the absorbance was measured [3, 50].
Oxidative stress and antioxidant defence
SOD, CAT, GSH, GSH-Px and GPx were investigated in the midbrain as previously described [51, 52] 24 hours (in the first experiment) or 30 days (in the second experiment) after the damage. ROS content was measured using commercial kits according to manufacturer guidelines [53].
Reagents and standards
The analytical standards of Gallic acid, Clorogenic acid, Catechin, Caffeic acid, syringic acid, rutin, ellagic acid, esperidin, ferulic acid, Miricetin, Quercetin, Apigenin, Naringenin, Kaempferol were purchased from Merck (Darmstadt, Germany). Acetonitrile and formic acid 99, 9% (LC-MS grade), water (HPLC gradient grade), were supplied from VWR (VWR International PBI Srl Milan, Italy). Acetone and HCl were purchased from Merck (Darmstadt, Germany). All the reagents were of HPLC grade (>99.9%).
Sample extraction and chromatographic analysis
The extraction of the samples was carried out according to the protocol proposed by Puigventós et al [54].. Briefly, 0.1 g of samples was mixed in 10 mL of acetone/water/hydrochloric acid (70:29.9:0.1 v/v/v) solution and sonicated for 30 min. Subsequently, the mixture was centrifuged at 3500 rpm at 15 min and stored at -4°C before the analysis. The chromatographic separation was carried out with a Raptor C18 column (100x2.1 mm, 2.7 µm) maintained at 25°C. The mobile phases consisted of H2O + 1% Formic acid (A) and acetonitrile + 1% Formic acid (B), with a 0.3 mL/min flow rate. The injection volume was 10 µL and all analytes are eluted with gradients from 5 to 95% A, for a total run of 13.63 min.
MS conditions and validation of the method
A Q Exactive™ Plus Hybrid Quadrupole-Orbitrap™ was used as mass spectrometer (Thermo Fisher Scientific, California, U.S.A.) with heated electrospray ionization source (HESI-II). The analysis was carried out in positive and negative polarity mode. All data were acquired using the Full MS scan/dd-MS2 mode. The operation parameters were according to D’Amico et al [27].: cover gas flow amount, 35 (random units); aux gas flow amount, 10 (random units); spray voltage, 3.50 kV; capillary temperature, 300 °C; tube lens voltage, 55 V; heater temperature, 305 °C; scan mode: full scan; scan range (m/z) 100–1000; microscans, 1 m/z; positive resolution: 70, 000; fourier transform (FT) automatic gain control (AGC) target: 3 × 10; negative resolution: 35, 000; automatic gain control (AGC) target: 1 × 106; maximum IT: 100 ms. The analytes were identified according to the retention time (tR), accurate mass and characteristic fragmentation. An external calibration for mass accuracy was performed each day before the analysis. Acquisition data were recorded and elaborated using Xcalibur TM version 4.0 software from Thermo.
The performance of the method was assessed in accordance with the Commission Decision 2002/657 for linearity, specificity, trueness. The LOQ was determined as the lowest amount of standard necessary to produce a S/N> 5 while being part of the linear range of the calibration curve (back-calculated residual < 20%). The linearity test gave satisfactory values for all the analytes examined (r2>0.996).
Materials
Unless otherwise stated, all compounds were purchased from Sigma-Aldrich.
Statistical evaluation
The data in this study are presented as the average ± SEM and represent at least three experiments conducted on various days. N denotes the number of animals utilized in in vivo experiments. The G*Power 3.1 software (Die Heinrich-Heine-Universität Düsseldorf, Düsseldorf, Germany) was employed to calculate the number of animals used in in vivo research. A competent histopathologist examined the data, without knowledge of the treatments. In all the statistical studies, GraphPad Software Prism 8 (La Jolla, CA, USA) was used. One-way or two way ANOVA was used to examine the data, and then a Bonferroni post-hoc test for multiple comparisons was used. A p value of 0.05 or less was regarded as significant.
Results
Sample extraction of HMPBE
In table 1 the results of the extraction of the samples of HMBPE.
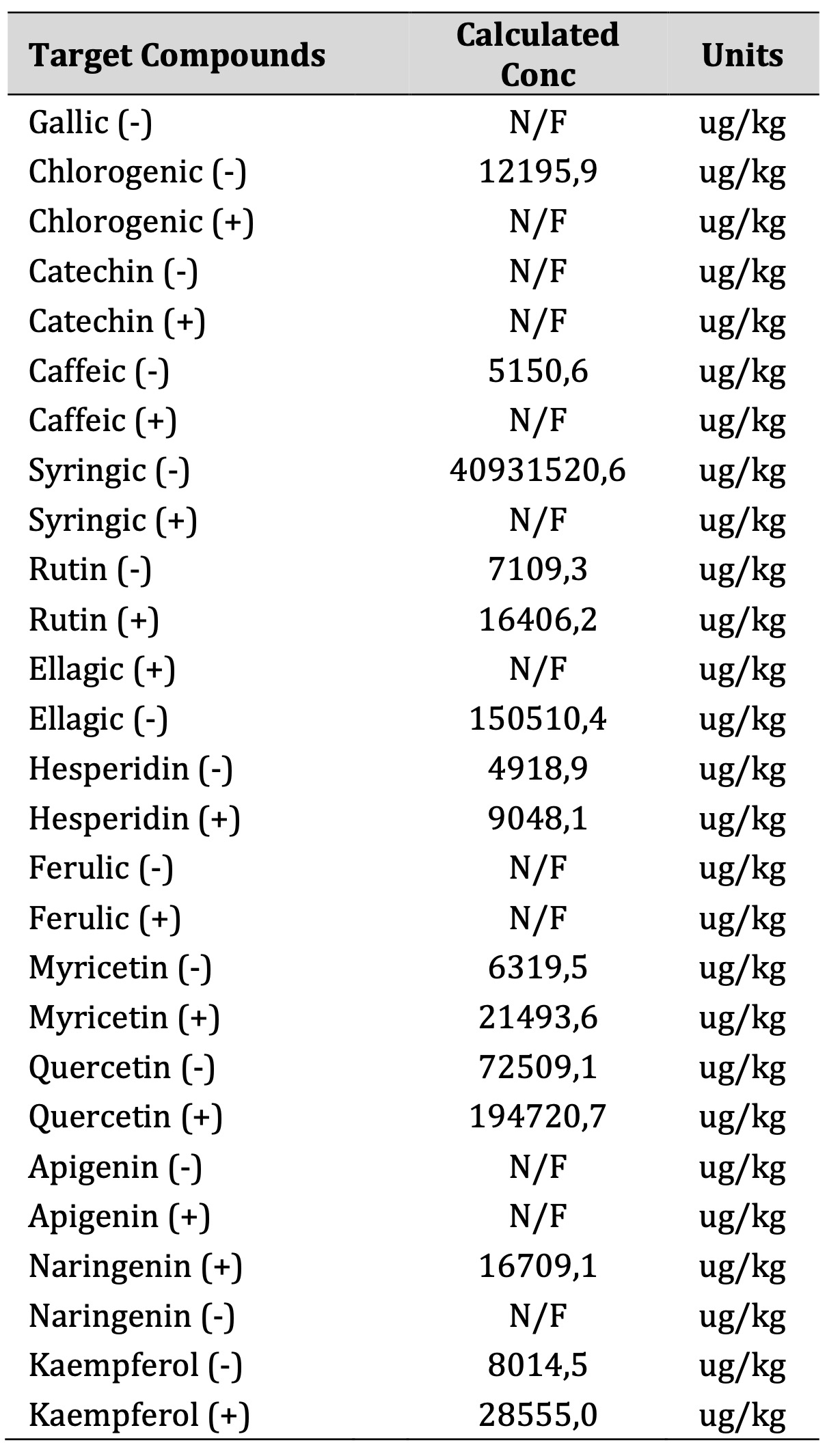
Table 1: chromatographic analysis HMBPE
Behavioural analysis of long-term admin-itrtion of HMPBE
To find the concentration for our studies, we chose to administered mice with HMPBE orally at three different increasing concentrations (5-15-30 Mg/kg) over a period of 3 months. Analysis of the data of Forced Swim (Fig. 1A), Open Field (Fig. 1B), Elevated Plus Maze (Fig. 1C), Morris Water Maze (Fig. 1D), Catalepsy (Fig. 1E), Rotarod (Fig. 1F) and pole (Fig. 1G) tests showing that the lowest concentration of HMPBE don’t have any significant effect on behaviour, compared to sham animals on the other hand 15 and 30 mg/kg improve significantly memory and spatial learning and ameliorate anxiety and depression. Additionally, during the first experiment performed, no cases of mortality, aggression, or significant changes in weight were observed.
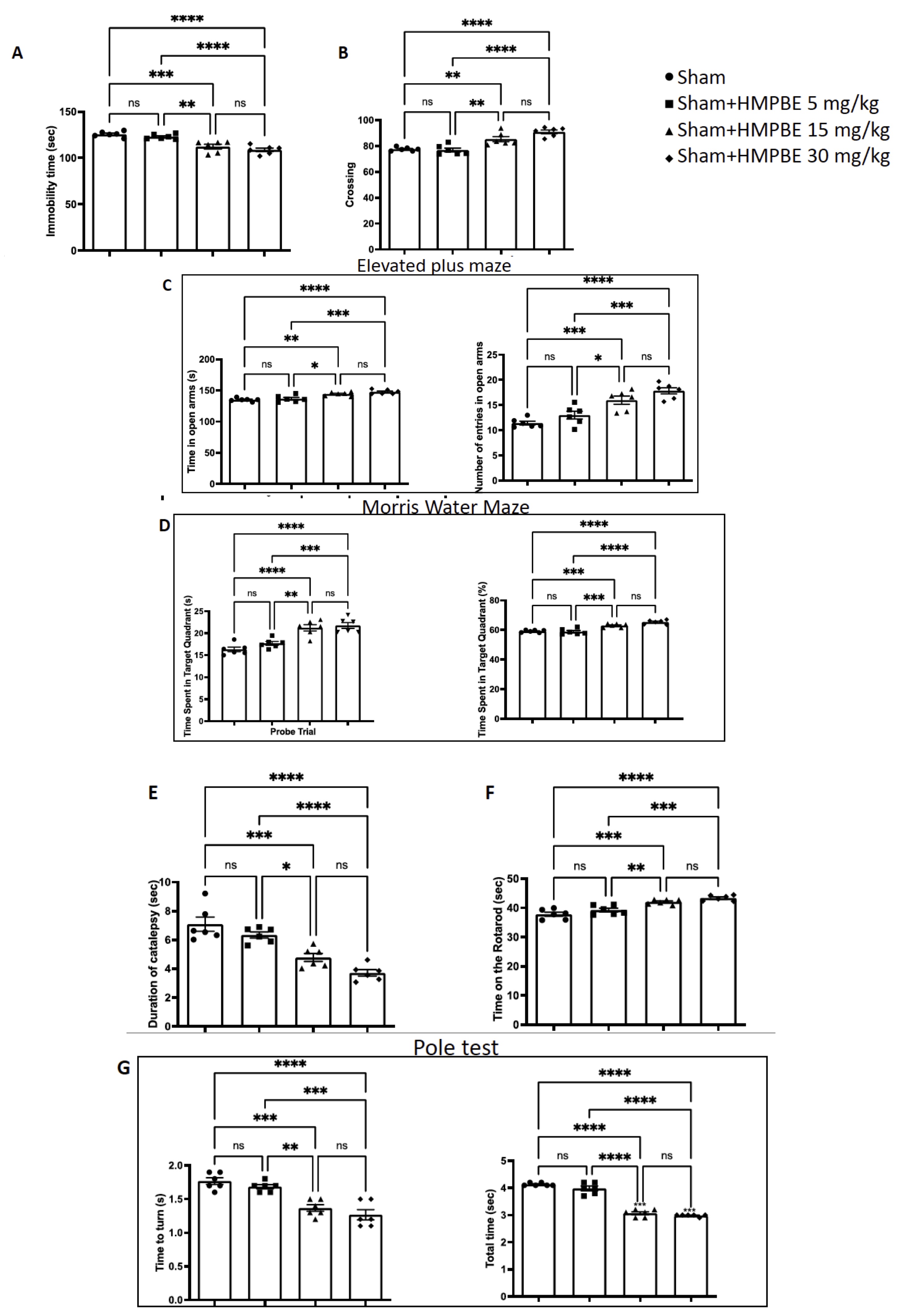
Fig. 1: Behavioural analysis of long-term administration of HMPBE. Forced Swim (A), Open Field (B), Elevated Plus Maze (C), Morris Water Maze (D), Catalepsy (E), Rotarod (F) and pole test (G) Statistical analysis made with GraphPad show the evaluation between all groups. *p <0.5; **p <0.01; ***p <0.001; ****P <0.0001. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group.
Effects of HMPBE on spatial learning, memory function post-injury anxiety and locomotor activity alterations induced by TBI
MWM test was performed to evaluate if HMPBE could have benefits on memory impairments induced by TBI. The time taken to find the platform during training was increased in TBI-subjected animals compared to the controls (Fig. 2A). In addition, the time spent in the target quadrant of the platform during probe trial was reduced in animal subjected to the trauma (Fig. 2A). Oral administration with HMPBE at the dose of 15 mg/kg significantly reduced the escape latency and increased the time spent in the target quadrant, indicating and improvement of the cognitive deficits induced by the trauma (Fig. 2A). We used the NOR (Fig. 2B) and SIT (Fig. 2C) tests to assess any deficits in their social interaction and exploratory behaviour. In both tests, we found that following TBI, exploration time spent on new or known items and the number of contacts were statistically higher (Fig. 2B and C, respectively), suggesting compromised cognitive function and a propensity to develop dementia-like symptoms. The administration of HMPBE, improve the recognition of familiar object but not of the novel (Fig. 2B). Additionally, significantly improve number of contacts in SIT (Fig. 2C). According to bibliography sham animals, spend more time in open arms, on the other hands, animals subjected to the injury spend more time in the closed arms (Fig. 2D), also reducing the number of entries (Fig. 2D). However, the animals that received the oral administration of HMPBE spent more time in the open arms (Fig. 2D) compared to TBI group also increasing the number of entries (Fig. 2D). The OFT was used to further evaluate locomotor activity. In contrast to sham mice, we discovered that after TBI injuries mice spent less time in the centre (Fig. 2E) also reducing the number of crossing (Fig. 2E). HMPBE was also successful in this instance in restoring locomotor activity (Fig. 2E) and the number of crossing (Fig. 2E).
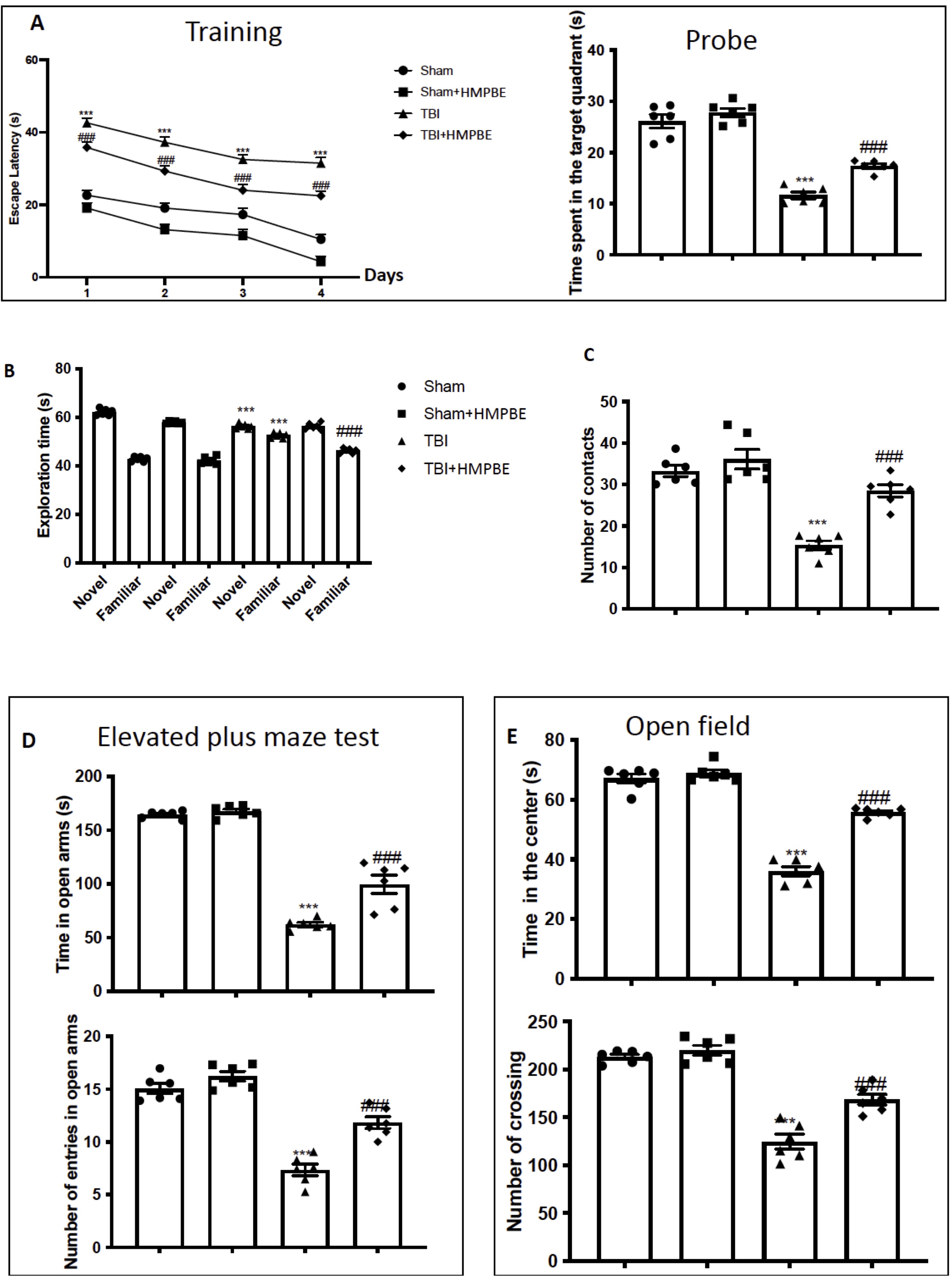
Fig. 2: Effects of HMPBE on spatial learning, memory function, anxiety, and locomotor activity. Morris Water Maze training and probe (A); novel object recognition (B); social interaction test (C); elevated plus maze test (D); Open field (E) ***p <0.001 vs sham; ###p <0.001 vs TBI. For the Fig. 2A statistical analysis was made with two way ANOVA. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group.
HMPBE limits histological alteration induced by TBI
When compared to brain from the sham group, histological investigation of sample at the level of the perilesional area revealed considerable tissue damage, inflammation, and architecture alterations in the TBI group 24 hours after TBI injury (Fig. 3A, A’ for sham and Fig. 3B, B’ for TBI, see histological score 3D). When compared to the TBI group, the HMPBE administration at the dose of 15mg/kg considerably lessen the severity of brain damage (Fig. 3C, C’ see histological score Fig. 3D).
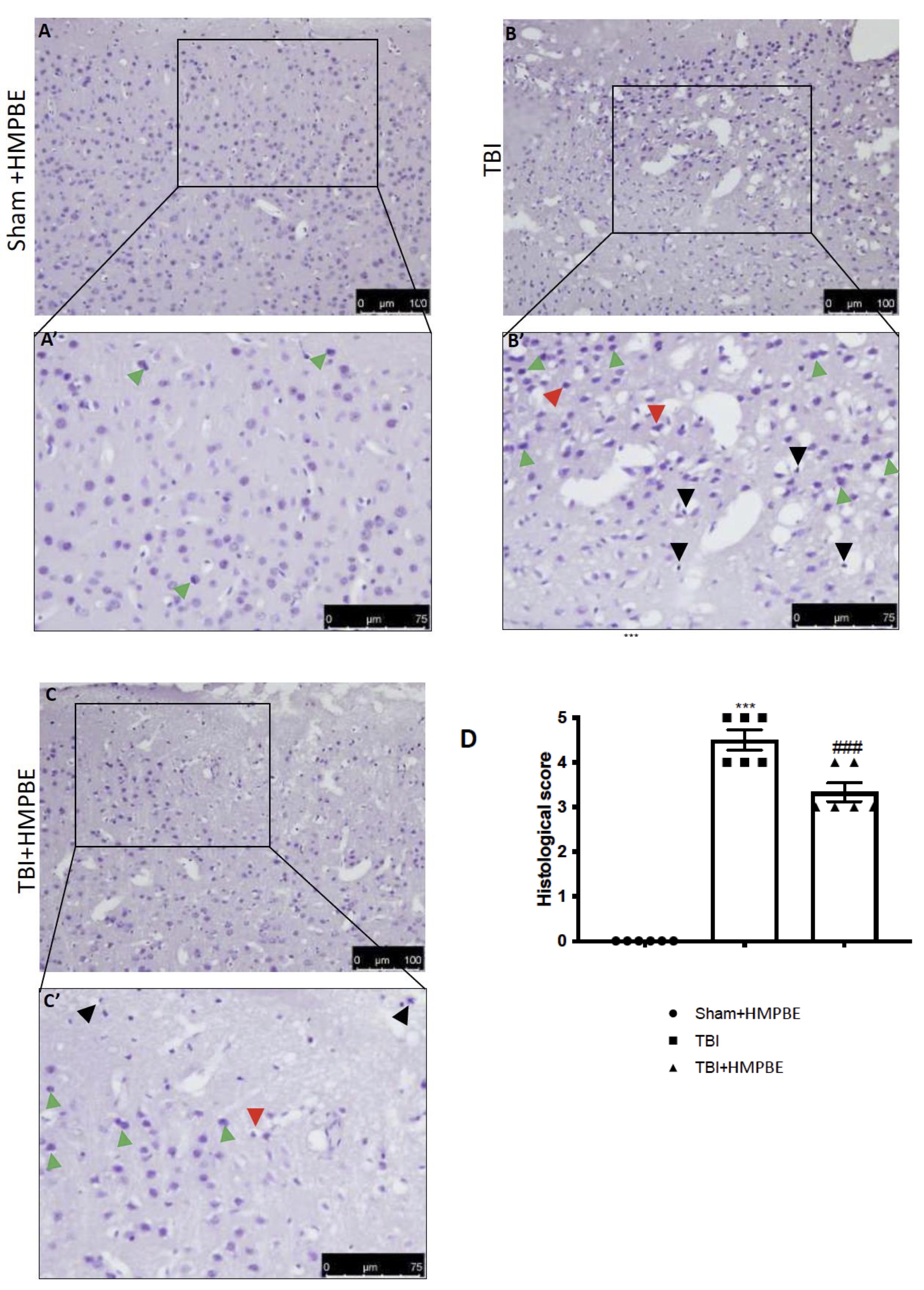
Fig. 3: HMPBE limits histological alteration induced by TBI. Representative images of histological structure of: Sham (A and higher magnification A’) TBI (B and higher magnification B’) and TBI+HMPBE (C and higher magnification C’). Red arrows indicates damaged neurons, green arrows indicates dark cells, black arrows indicates eosinophilic infiltration. Histological score (D) **p <0.01vs sham; ***p <0.001 vs sham; ##p <0.01 vs TBI; ###p <0.001 vs TBI. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group.
Effects of HMPBE administration on inflammatory and antioxidant pathways
To further investigated the molecular pathways by which HMPBE could act, we analysed by western blots two well-know inflammatory and anti-oxidative players involved in the secondary damage TBI-related in the first 24 hours after the injury. According to bibliography we found that after TBI there were a significantly decrease of Ikb-a (Fig. 4A) and as consequence a significantly increase in nf-kb translocation (Fig. 4B) in the animals subjected to TBI compared to the sham group. Additionally, we found and increase in in Nrf-2 activation (Fig. 4D) and consequently in HO-1 expression (Fig. 4C), compared to the control group. HMPBE administration at the dose of 15 mg/kg was able to restore nf-kb translocation (Fig. 4B) as well as Ikb-a (Fig. 4A) almost at the physiological levels and additionally was able to significantly improved Nrf-2 activation (Fig. 4D) and consequently in HO-1 expression (Fig. 4C). The improving of the physiological antioxidant endogenous system was also reflected in the increasing of SOD (figure 4E) and GSH-Px (Fig. 4F) activity obtained after HMPBE administration.
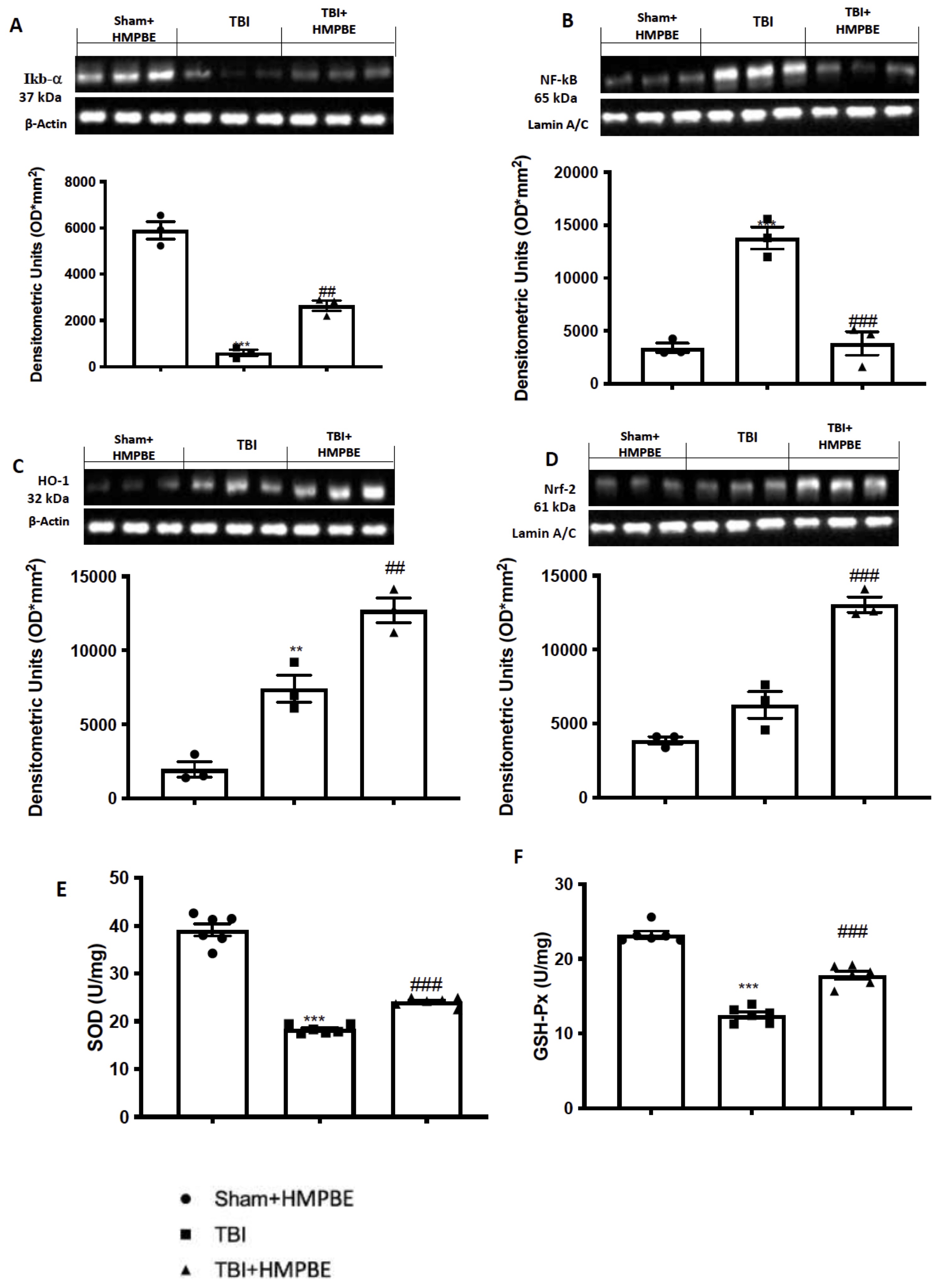
Fig. 4: Effects of HMPBE administration on inflammatory and antioxidant pathways. Representative images of western blots of and relative quantification: Ikb- (A), Nf-kb (B), HO-1 (C) and Nrf-2 (D). SOD (E) and GSH-Px activity (F). ***p <0.001 vs sham; ###p <0.001 vs TBI. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group
Effects of HMPBE on apoptosis, lipid peroxidation and cytokines release
According to reports, 24 hours after the injury TBI triggered apoptotic pathways. For this reason, we identified the expression levels of the proteins Bax (Fig. 5A) and Bcl-2 (Fig. 5B), two elements of the mitochondrial mechanism of death that are antagonistic to one another. After TBI we notice a significantly increase in Bax expression and vice versa a significantly decrease in Bcl-2 expression. Given the large concentration of polyunsaturated fatty acids in the brain, oxidative stress after TBI primarily appears as lipid peroxidation. We found that in TBI group there was a significantly increase in lipid peroxidation (Fig. 5C), compared to sham animals. Moreover, since cytokines are among the additional downstream gene targets of Nf-kb, we used ELISA kits to measure the levels of TNF-a (Fig. 5D), IL-6 (Fig. 5E), and IL-1b (Fig. 5F). While low amounts of this cytokine were identified in the sham group, brain tissues taken from TBI mice showed a significant rise in all cytokines. HMPBE administration at the dose of 15 mg/kg significantly reduce apoptosis as well as lipid peroxidation and cytokines release.
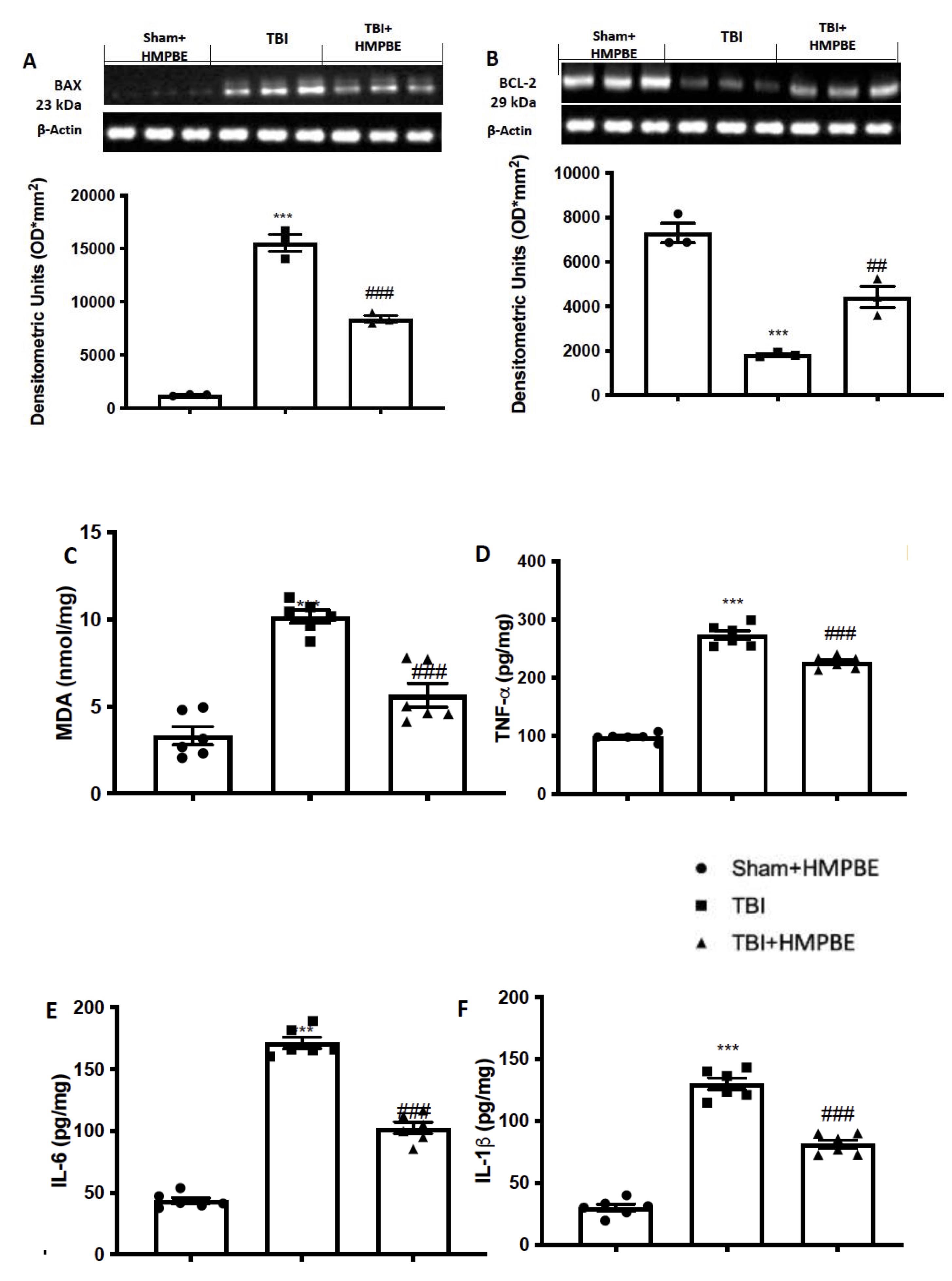
Fig. 5: HMPBE reduced apoptosis, lipid peroxidation and cytokines release. Representative images of western blots and relative quantification of Bax (A) and Bcl-2 (B); MDA levels (C); ELISA quantification of TNF-a (D), IL-6 (E), and IL-1b (F). ***p <0.001 vs sham; ##p <0.01 vs TBI; ###p <0.001 vs TBI. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group
Effects of HMPBE administration on long term behavioural alteration and histological damage induced by TBI
As performed previously in acute, we compared the level of anxiety and locomotor function in mice who had chronic TBI using EPM and OF tests. We noticed that TBI animals spent less time in open arms during the EPM test than control mice (Fig. 6A) as well as a decrease numbers of crossing (Fig. 6B). Moreover, OF test further supported the anxiety level seen in mice with the injury. Compared to Sham animals, mice with chronic TBI displayed more thigmotaxis, and a propensity to remain near the wall or outside the field (Fig. 6C and 6D). Animals that received daily oral administration with HMPBE at the dose of 15 mg/kg exhibited behaviour that was like that of the Sham group; in fact, they were found to learn to approach the black escape box and exit the open field with a considerable reduction in escape latency. 30 days after the impact, histological analysis of the brains taken from TBI animals (Fig. 6F and F’, see histological score 6H) revealed considerable tissue modification in the cortex’s perilesional area as compared to Sham mice (Fig. 6E and E’, see histological score 6H). Here, we found that HMPBE decreased the severity of brain damage accelerating the physiological recovery (Fig. 6G and G’, see histological score 6H).
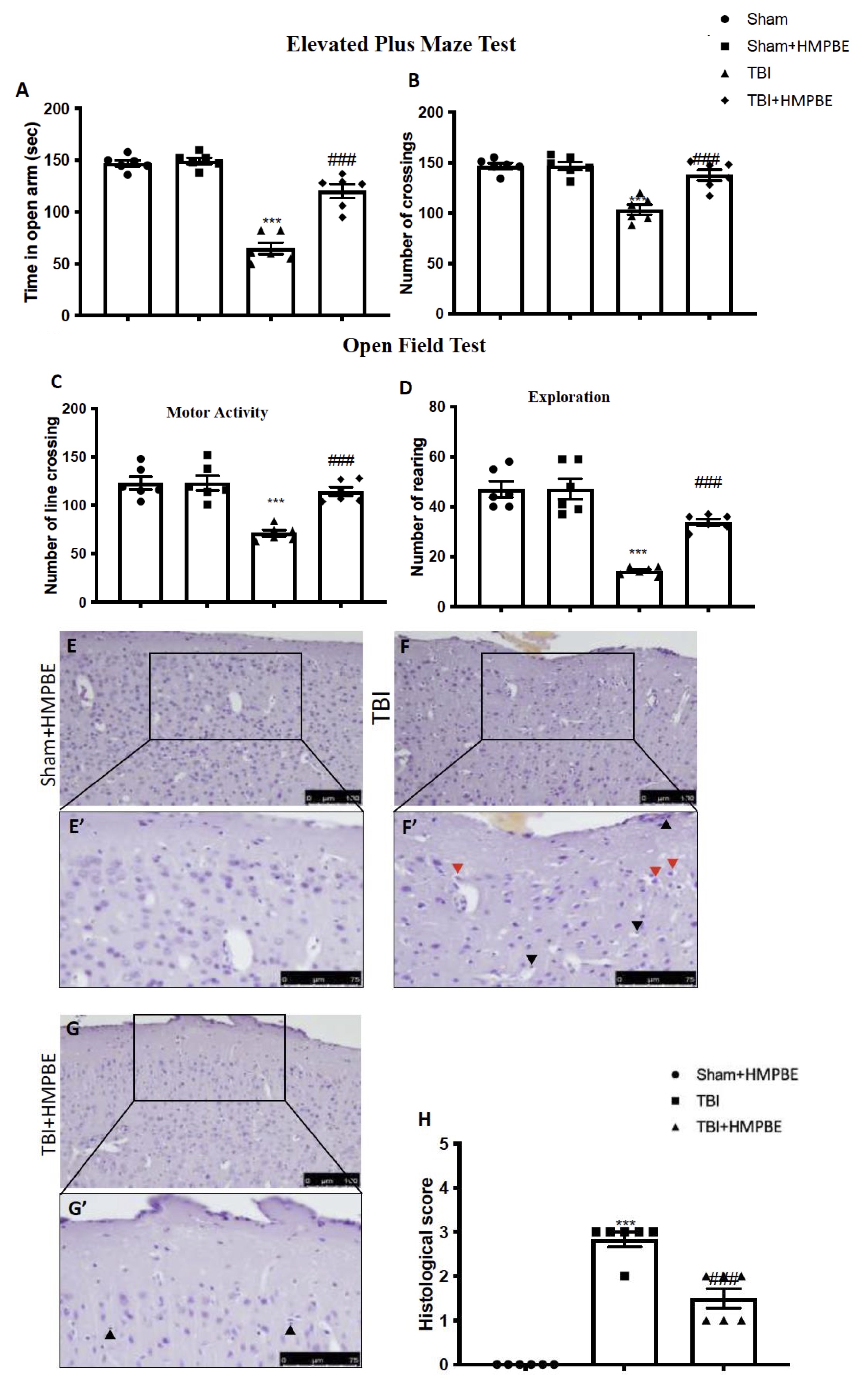
Fig. 6: HMPBE ameliorate post-injury anxiety, locomotor activity and improve histological recovery after chronic TBI. Elevated Plus Maze test (A) and (B); Open field test (C) and (D); representative images of histological structure of: Sham (E and higher magnification E’) TBI (F and higher magnification F’) and TBI+HMPBE (G and higher magnification G’). Histological score (H). Red arrows indicates damaged neurons, black arrows indicates eosinophilic infiltration. ***p <0.001 vs sham; ###p <0.001 vs TBI. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group.
HMPBE administration reduced Nf-kb pathways activation
We also used Western blot analysis to examine the expression of Nf-kb pathways to determine how HMPBE reduced the neuroinflammatory processes that resulted in the development of PD following TBI. The results revealed that Ikb-a(Fig. 7A) was expressed at a low level in the control group while a large amount of this protein’s breakdown was seen in response to TBI. According to our findings, levels of Nf-kb nuclear translocation (Fig. 7B) were low in the Sham group and significantly higher 30 days after TBI. Additionally, we investigated the same parameters taken in consideration before, during chronic TBI. We notice that 30 days after TBI there were an increase in increase in MDA (Fig. 7C) levels as well as in TNF-a (Fig. 7D), IL-1b (Fig. 7E), and IL-6 (Fig. 7F). As showed after acute TBI, also in this case, HMPBE administration significantly reduce the activation of Nf-kb pathways as well as lipid peroxidation and cytokines release.
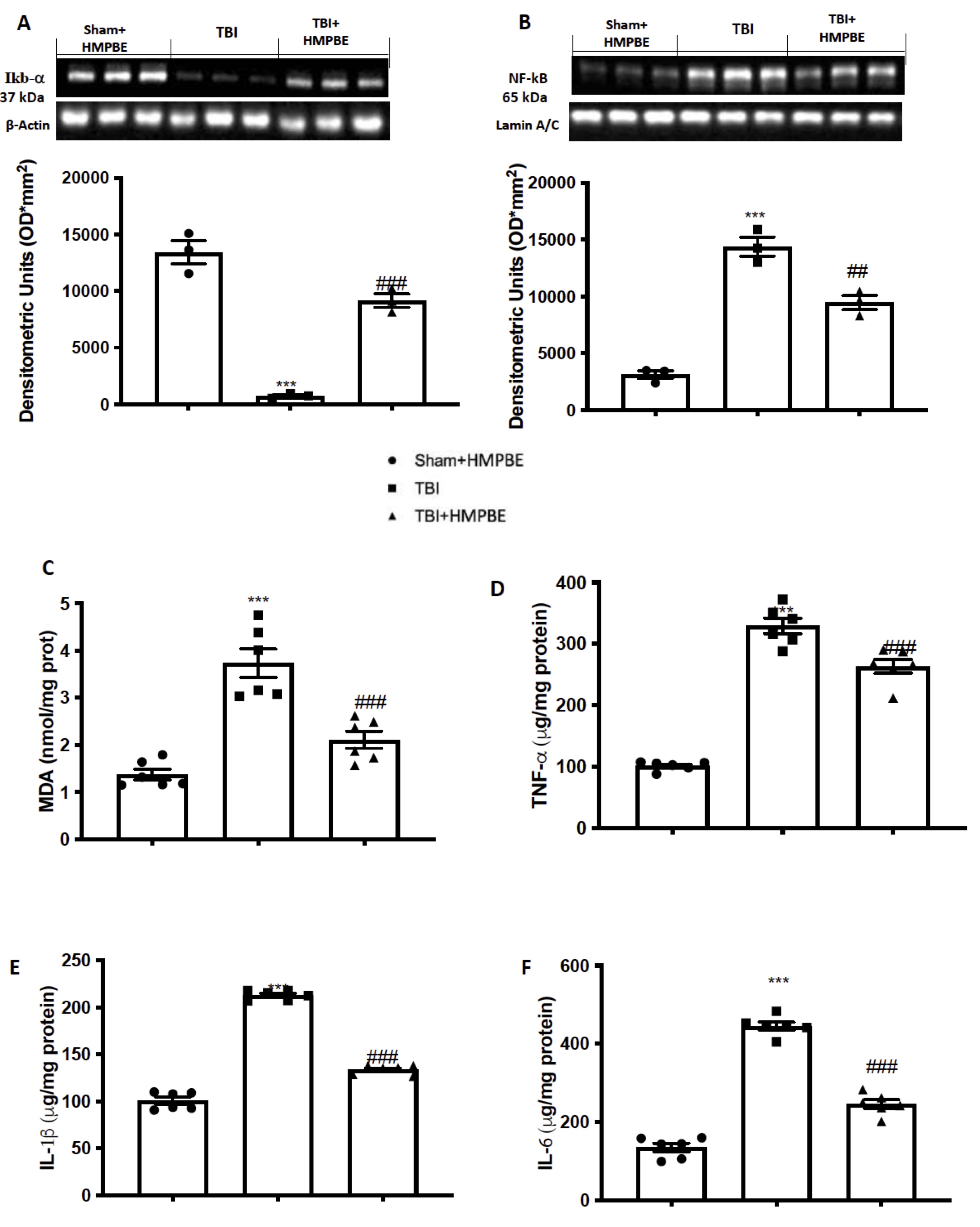
Fig. 7: HMPBE reduced Nf-kb activation, lipid peroxidation and cytokines release after chronic TBI. Representative images of western blots and relative quantification of: Ikb- (A), Nf-kb (B), MDA levels (C); ELISA quantification of TNF-a (D), IL-1b (E), and IL-6 (F). ***p <0.001 vs sham; ##p <0.01 vs TBI; ###p <0.001 vs TBI. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group
Influence of HMPBE on oxidative stress after chronic TBI
Increased production of genes that aid in the removal of free radicals, the restoration of mitochondrial function, and genes sensitive to cell survival stress, known as vitagenes, are all linked to oxidative stress in the brain that arises after TBI [27]. In this context, we aimed to assess how the Nrf2/HO-1 pathway was affected by HMPBE administration. Our results demonstrated that 30 days after TBI induction there were a significantly increase in brain ROS content (Fig. 8A) and a physiological activation of Nrf-2 pathways (Fig. 8B) strongly connected with HO-1 expression (Fig. 8C). Furthermore, a basal Nrf-2 signal can be seen in the sham animal sections (Figures 2S, panel 2D), which physiologically increases in animals subjected to TBI (Figures 2S, panel 2E), which following administration with HMBPE is significantly increased (Fig. 2S, panel 2F).On the other hand, 1 months after the trauma, physiological antioxidant defence in significantly decrease as demonstrated by the reduction of SOD (Fig. 8D), CAT (Fig. 8E), GPx (Fig. 8F) and GSH (Fig. 8G) activity. Daily administration of HMPBE significantly improve the translocation of Nrf-2, increasing the endogenous antioxidant system and reducing oxidative stress in the brain.
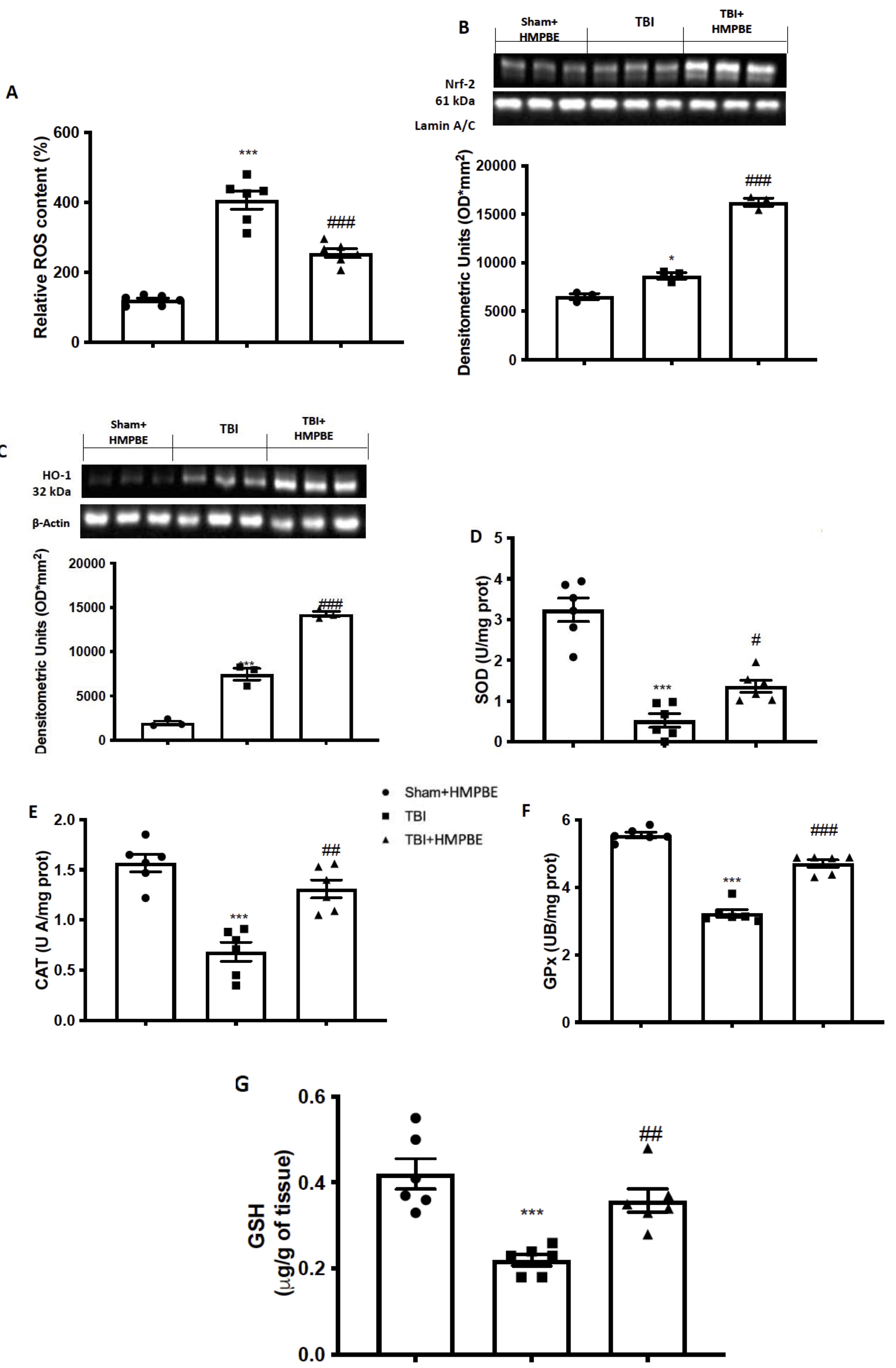
Fig. 8: Effects of daily administration of HMPBE against oxidative stress induced by chronic TBI. ROS content (A); representative images of western blots and relative quantification of Nrf-2 (B) and HO-1 (C); SOD (D); CAT (E); GPx (F) and GSH (G). *p <0.05 vs sham ***p <0.001 vs sham; #p <0.05 vs TBI; ##p <0.01 vs TBI ###p <0.001 vs TBI. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group.
HMPBE administration’s impact on PD Marker changes during chronic TBI
Midbrain slices from mice in each group were stained with TH to see if chronic TBI could change PD-like marker. When compared to the Sham group (Fig. 9A), the expression of TH-positive neurons was significantly reduced 30 days after TBI (Fig. 9B). a-syn and DAT, two crucial PD biomarkers, were also processed by western blots in midbrain areas. We found that compared to the Sham group, chronic TBI clearly increased a-syn (Fig. 9E) and significantly decreased DAT expression (Fig. 9F). HMPBE was given daily at a dose of 15 mg/kg, and we observed a considerable restoration of TH positive neurons (Fig. 9C) and DAT expression (Fig. 9F) as well as a reduction in a-syn accumulation (Fig. 9F).
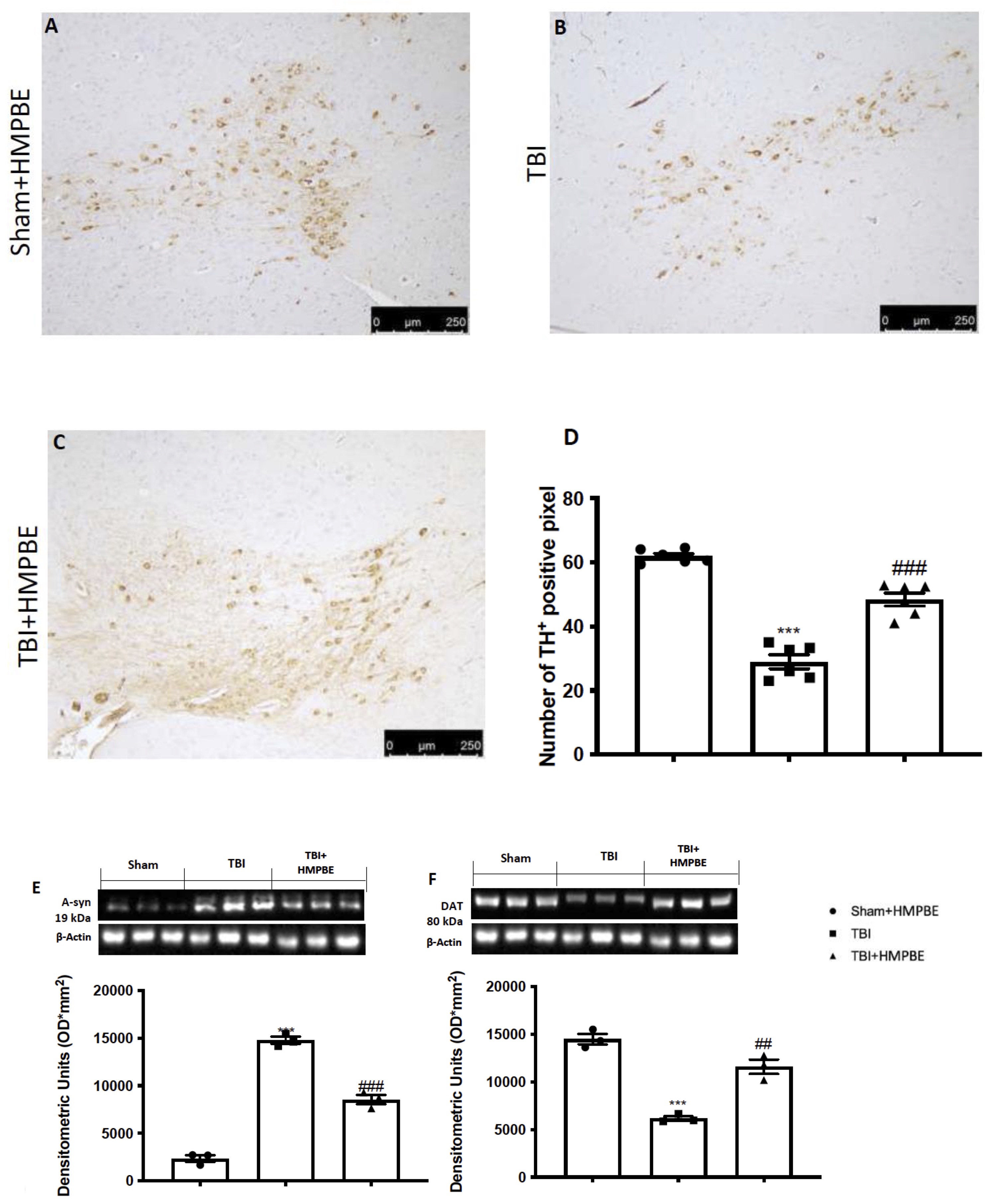
Fig. 9: HMPBE administration's impact on PD Marker changes during chronic TBI. Immunohistochemical analysis of TH in midbrain section from: Sham (A), TBI (B) and TBI+HMPBE (C); quantification of TH positive neurons (D). Western Blot analysis and relative quantification of a-syn (D) and DAT (F). ***p <0.001 vs sham; **p <0.01 vs sham; #p <0.05 vs CYP; ##p <0.01 vs TBI; ###p <0.001 vs TBI. The Fig.s are representative of at least three experiments performed on different experimental days. Each data are expressed as mean±S.E.M. from n=6 male mice for each group.
Discussion
One of the most significant public health problems is TBI, which is commonly referred to as a “silent epidemic” since the difficulties it causes take time to manifest. The processes behind the TBI pathology have been considerably defined by research. These involve a primary injury brought on by direct biomechanical forces and a secondary injury that follows a traumatic brain injury and causes brain damage and death. A TBI may cause behavioural and physical symptoms, and it may leave the victim completely recovered or permanently disabled. Additionally, TBI-related secondary events can affect the patients’ quality of life. Patients may have cognitive deterioration, dementia risk, and brain atrophy [6]. First reports of Nrf2’s neuroprotective effects on TBI date back to 2007[55]. Numerous investigations have subsequently shown that Nrf2 protected against TBI. In recent decades, Nrf2 has drawn more attention as a crucial part of cellular defence against a range of endogenous and exogenous stressors [56]. Numerous studies show that Nrf2 functions as a downstream target of therapeutic drugs in addition to being a significant protective element in damage models. Numerous studies have determined that Nrf2 activation influences TBI by preventing oxidative stress, apoptosis, and inflammation through a variety of molecules and pathways, including HO-1, Bcl-2, and NF-κB [57, 58]. More than 200 cytoprotective proteins are regulated by Nrf2, which is specifically the master regulator of antioxidant defences in response to oxidative stress. Under ordinary conditions, Nrf2 is anchored by its inhibitor Keap1 and is found in the cytoplasm. On the other hand, Nrf2 separates from Keap1 in the presence of oxidative or xenobiotic stress, moves into the nuclear region, and interacts with the ARE to regulate the expression of antioxidant genes. Furthermore, it is possible that Nrf2 activation could lessen the secondary damage that results from a TBI because oxidative stress brought on by the generation of ROS causes significant damage to brain tissue [59]. Apoptosis in TBI exacerbates damage and shortens survival time by causing additional neuronal death. Researchers have investigated the potential impact of Nrf2 on TBI-induced apoptosis in this regard. According to Jin et al.’s findings, mice lacking Nrf2 had higher rates of neuronal death in their brains [60]. The precise nature of the connection between Nrf2 and NF-κB remains unclear; nevertheless, the discovery of NF-κB binding sites in the Nrf2 gene’s promoter region raises the possibility of cross-talk between these two regulators during inflammatory processes. It has been suggested that Keap1 inhibits IKK-β phosphorylation and mediates IKK-β degradation by autophagocytosis, thereby acting as a negative regulator of NF-κB signaling. Each of these investigations indicated that, depending on the circumstance, Nrf-2, Bcl-2, and NF-κB interacted with one another. Nonetheless, there have been reports of both positive and negative regulation between Nrf-2, Bcl-2, and NF-κB; hence, more research is necessary to understand the interplay among these pathways [61-63]. Numerous phytochemicals have recently been discovered in natural plants, including fruits, vegetables, grains, and other therapeutic herbs, and they have been found to downregulate nf-kb or to stimulate the Nrf2 signalling pathway to have neuroprotective effects in TBI [64]. In detail, about nf-kb, it was previously demonstrated that after being transported into the cells, monomers, dimers, and related metabolites can act in the following ways: (1) reducing the rise in oxidants linked to certain stimuli and the subsequent activation of NF-KB (antioxidant effect); (2) blocking IKK-related events upstream or directly inhibiting IKK; (3) preventing the phosphorylation and/or degradation of the inhibitory protein IKBa; (4) interfering with the movement of active NF-B from the cytosol into the nucleus; (5) preventing NF-kB from binding to NF-KB DNA sites, either directly through interactions with NF-kB proteins or by interacting with DNA. Large procyanidins (three units or more) that are not taken up by the cells may be able to control the membrane-level activation of NF-kB. Procyanidins have been shown to interact with both synthetic and biological membranes, changing the physical characteristics of the membranes. Therefore, flavanols and procyanidins may either (1) alter the biophysical properties of the membrane and alter the receptor environment, or (2) directly interact with the receptor to influence the binding of the ligand (stimuli) to its receptor [65-68]. On the other hand, these organic substances act on Nrf-2 pathways thanks to the ability to: (i) oxidize or alkylate Keap-1 proteins; (ii) prevent Nrf2 from attaching to Keap-1; and (iii) phosphorylate Nrf2 by GSK-3, which leads to proteasomal destruction [69, 70]. Studies on HMPBE are still few and limited, for this reason, in our first step we wanted to demonstrate that the prolonged administration of HMPBE at different concentrations was able to improve mice behaviour. Using 3 different doses we established that the lowest dose with a significant effect was 15 mg/kg. After identifying the minimum dose with a beneficial effect on behaviour, we used this dose to plan the next two studies, acute and chronic TBI. Animal models of trauma are frequently used to study the prevalent clinical issue in humans. The acute inflammatory response causes nuclear transcriptional factor activation, pro-inflammatory complex assembly, release of inflammation mediators, and an increase in oxidative stress after regulated cortical impact [24, 25]. TBI also causes secondary processes in animals that result in neurological conditions like impaired memory and learning as well as seizures. In our study, a single oral administration 1 h after trauma induction was able to improve spatial learning and memory function as well as post-injury anxiety and locomotor activity altered after the trauma. The molecular pathways by which its act it’s probably due to its ability to modulate Nf-kb as well as Nrf-2 and apoptotic pathways. According to the literature, by modulating these two pathways the entire downstream cascade was also modulated including cytokine release and lipid peroxidation, as well as enhancement of physiological antioxidant enzymes. The physiological responses to PD and TBI are comparable, therefore it’s likely that TBI could accelerate underlying PD pathology in an additive or synergistic way, causing subclinical PD disease to become overt PD pathology. Since both mice and rats have physiological responses that are comparable to those found in post-mortem human brain tissue, these animals make an excellent study platform for future studies that will clarify how inflammation may influence TBI pathophysiology and lead to the development of PD [71, 72]. Even 30 days after TBI the neuroinflammatory and oxidative stress events were still present in the brain together with the reduction of dopaminergic neurons and the accumulation of a-syn protein. Impellizzeri et al. in particular demonstrated that the TBI inflicted on mice after CCI results in a considerable decline in PD-specific markers like TH and DAT after 30 days and in an accumulation of a-syn into midbrain region [73]. In this third experiment, daily administration of HMPBE, at the dose of 15 mg/kg was able to reduce histological alteration as well as inflammation and lipid peroxidation. Additionally, it was able to significantly improve behavioural recovery as well as physiological antioxidant response. Last but not least, HMPBE has been shown to be able to reduce the alterations that from TBI lead to PD by limiting the death of dopaminergic neurons, increasing the expression of DAT and reducing the accumulation of a-syn [74]. In conclusion, HMPBE has been demonstrated for the first time to be able to act on molecular processes involved in acute and chronic brain trauma. The neuroinflammation of the brain appears to be reduced in part because of its apparent action on the Nf-kb pathway. Additionally, this compound’s effects on oxidative stress are accompanied by an increase in the physiological antioxidant response due to the modulation of the Nrf-2 pathway. Chronic neuroinflammation has come to light as a potential contributory element to behaviour change along with these cell stress responses. Preclinical studies have demonstrated that a large inflammatory load is linked to TBI. Additionally, it has been demonstrated that neuroinflammation in the brains of retired sportsmen can last for years after an injury. Neuroinflammation and neurobehavioral symptoms are linked by recent clinical research. We assume that reducing cellular stress after TBI will have a favourable impact on the extracellular inflammatory environment, leading to better behavioural results [75, 76]. For this reason, in addition to the results demonstrating the beneficial effects of HMPBE administration on learning and memory processes, we demonstrated that it was able to improve the behavioural alterations induced by trauma both during the acute and chronic phases, limiting thus the onset of anxiety to depression. HMPBE may therefore be utilized as dietary supplements to stop the neurodegenerative effects of chronic traumatic brain injury (Fig. 10).
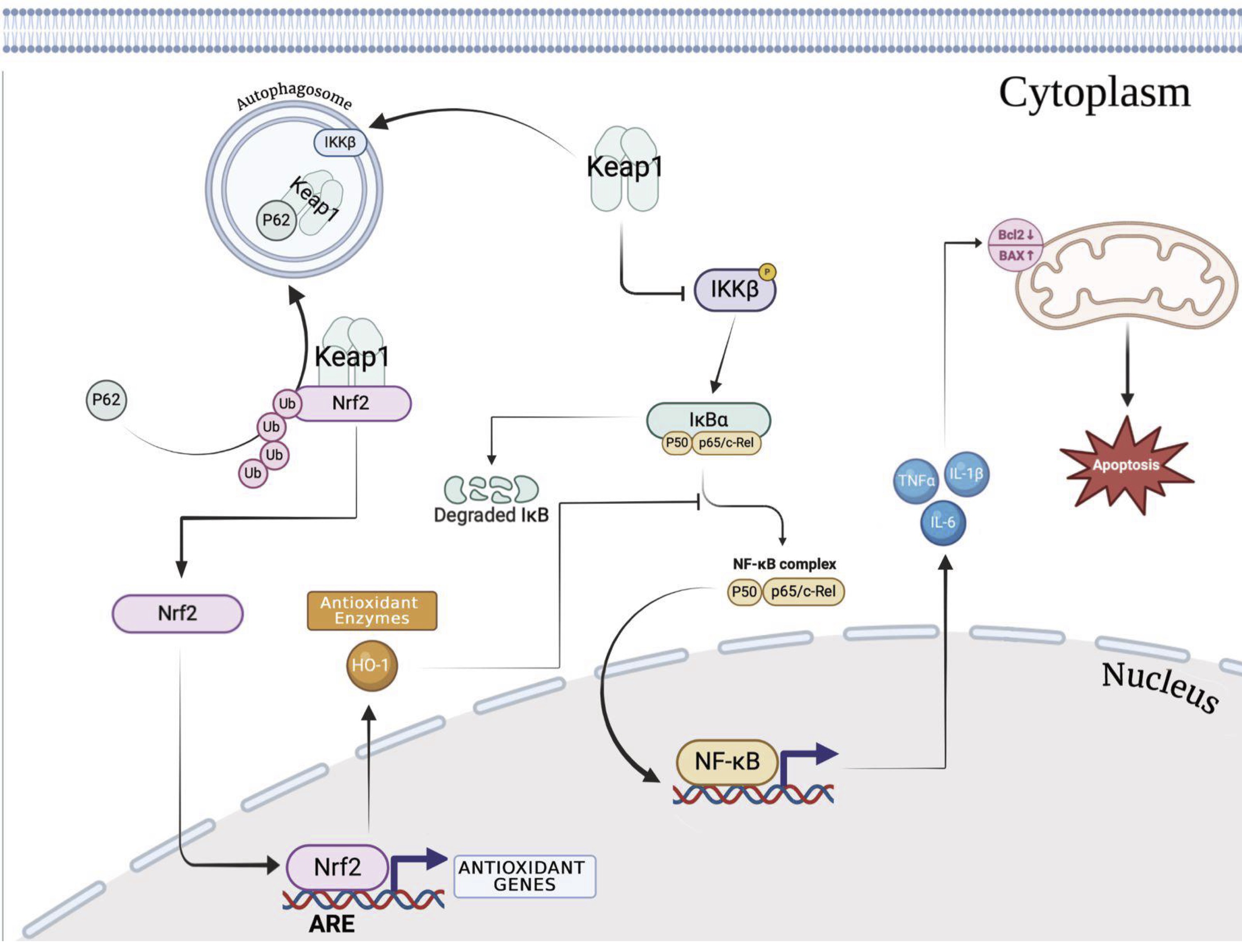
Fig. 10: Pathways involved and regulated by HMBPE in acute and chronic brain injury. Schematic representation on pathways involved in acute and chronic TBI and mechanism of action of HMBPE
Conclusion
This study provides the first evidence that HMPBE can counteract inflammation, oxidative stress and apoptosis induced after TBI in the mouse in both acute and chronic. Given that neuroinflammation plays an important role in neurotrauma, the investigation of free radical generation, as well as determining the antioxidant abilities and the effects of antioxidants, may be important for evaluating the overall balance of oxidative stress after acute and chronic brain injury. Moreover, in this study we demonstrate for the first time that HMPBE administration could be helpful to ameliorate cognitive deficits induced after the trauma. Acute neuroprotective therapies aim to block the molecular cascade of injury following TBI. Although neuroprotection is an important strategy for the treatment of such injury, to date, no effective neuroprotective agents have been identified from TBI clinical trials. Another important aspect is that most clinical trial strategies have used compounds that target a single pathophysiological mechanism, even though many mechanisms are involved in secondary injury after TBI especially during long term recovery. Our study possesses many limitations related to the other TBI-induced event such as has reactive astrogliosis, neuron loss, axonal damage, demyelination, and microgliosis, which have an effect on the entire recovery. Additionally, more studies are mandatory to pharmacokinetics, bioavailability, mood of action of these components and all the cascade events it begins with the impact and lasts throughout the patient’s life. Future prospecting of our study could be testing HMBPE on different aspects and consequences of trauma that have not been taken into consideration to date, such as double the risk after a trauma of suicide and is a primary determinant for the development of acquired seizure disorders, significantly increases in the risk of AD, and chronic traumatic encephalopathy (CTE) to investigate whether the unique combination of this compound is able to act on multiple fronts.
Acknowledgements
Ethics approval
The University of Messina Review Board for animal care (OPBA) approved the study. All animal experiments agree with the new Italian regulations (D.Lgs 2014/26), EU regulations (EU Directive 2010/63) and the ARRIVE guidelines.
Availability of data and materials
The datasets generated and/or analyzed for the present study are available from the corresponding author on reasonable request.
Funding
This research has been conducted with funds from MUR PRIN 2022FWB4EX, University of Catania, Italy and funds from Research Innovation, Ministry of Education in Saudi Arabia Project number RSPD2024R750, King Saud University, Riyadh, Saudi Arabia.
Authors’ contributions
Rosanna Di Paola and Marika Cordaro: Conceptualization, Writing - review & editing; Ramona D’Amico, Ali S. Abdelhameed and Uwe Wenzel; Data curation, Formal analysis; Livia Interdonato, Roberta Fusco, Daniela Impellizzeri, Rosalba Siracusa: Investigation, Methodology; Salvatore Cuzzocrea and Vittorio Calabrese: Project administration, Funding acquisition; Ursula Jacobs, Tilman Fritsch and Naomi Osakabe: Supervision, Validation; Sergio Modafferi and Francesco Molinari: Roles/Writing - original draft. All authors approved the version to be published and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Disclosure Statement
The authors have no relevant financial or non-financial interests to disclose.References
| 1 | Meaney DF, Morrison B, Dale Bass C: The mechanics of traumatic brain injury: a review of what we know and what we need to know for reducing its societal burden. J Biomech Eng 2014;136(2):021008. DOI: 10.1115/1.4026364.
https://doi.org/10.1115/1.4026364 |
| 2 | Huang X, Guo X, Gao W, Xiong Y, Chen C, Zheng H, Pan Z, Wang L, Zheng S, Ke C, Stavrinou P, Hu W, Hong K, Zheng F: Causal association between years of schooling and the risk of traumatic brain injury: A two-sample mendelian randomization analysis. J Affect Disord 2024;354:483-90. DOI: 10.1016/j.jad.2024.03.045.
https://doi.org/10.1016/j.jad.2024.03.045 |
| 3 | Genovese T, Impellizzeri D, D'Amico R, Fusco R, Peritore AF, Di Paola D, Interdonato L, Gugliandolo E, Crupi R, Di Paola R, Cuzzocrea S, Cordaro M, Siracusa R: Role of Bevacizumab on Vascular Endothelial Growth Factor in Apolipoprotein E Deficient Mice after Traumatic Brain Injury. Int J Mol Sci 2022;23(8). DOI: 10.3390/ijms23084162.
https://doi.org/10.3390/ijms23084162 |
| 4 | Campolo M, Casili G, Lanza M, Filippone A, Cordaro M, Ardizzone A, Scuderi SA, Cuzzocrea S, Esposito E, Paterniti I: The inhibition of mammalian target of rapamycin (mTOR) in improving inflammatory response after traumatic brain injury. J Cell Mol Med 2021;25(16):7855-66. DOI: 10.1111/jcmm.16702.
https://doi.org/10.1111/jcmm.16702 |
| 5 | Padmakumar S, Kulkarni P, Ferris CF, Bleier BS, Amiji MM: Traumatic brain injury and the development of parkinsonism: Understanding pathophysiology, animal models, and therapeutic targets. Biomedicine & Pharmacotherapy 2022;149:112812. DOI: https://doi.org/10.1016/j.biopha.2022.112812.
https://doi.org/10.1016/j.biopha.2022.112812 |
| 6 | Delic V, Beck KD, Pang KCH, Citron BA: Biological links between traumatic brain injury and Parkinson's disease. Acta Neuropathol Commun 2020;8(1):45. DOI: 10.1186/s40478-020-00924-7.
https://doi.org/10.1186/s40478-020-00924-7 |
| 7 | Sharma A, Muresanu DF, Nozari A, Lafuente JV, Sahib S, Tian ZR, Buzoianu AD, Patnaik R, Wiklund L, Sharma HS. Chapter 4 - Mild traumatic brain injury exacerbates Parkinson's disease induced hemeoxygenase-2 expression and brain pathology: Neuroprotective effects of co-administration of TiO2 nanowired mesenchymal stem cells and cerebrolysin. In: Sharma HS, Sharma A, editors. Progress in Brain Research. Elsevier; 2020. p. 157-231.
https://doi.org/10.1016/bs.pbr.2020.09.010 |
| 8 | Ulusoy A, Di Monte DA: α-Synuclein elevation in human neurodegenerative diseases: experimental, pathogenetic, and therapeutic implications. Mol Neurobiol 2013;47(2):484-94. DOI: 10.1007/s12035-012-8329-y.
https://doi.org/10.1007/s12035-012-8329-y |
| 9 | Huang M, Wang B, Li X, Fu C, Wang C, Kang X: α-Synuclein: A Multifunctional Player in Exocytosis, Endocytosis, and Vesicle Recycling. Front Neurosci 2019;13:28. DOI: 10.3389/fnins.2019.00028.
https://doi.org/10.3389/fnins.2019.00028 |
| 10 | Lashuel HA, Overk CR, Oueslati A, Masliah E: The many faces of α-synuclein: from structure and toxicity to therapeutic target. Nat Rev Neurosci 2013;14(1):38-48. DOI: 10.1038/nrn3406.
https://doi.org/10.1038/nrn3406 |
| 11 | Barnham KJ, Masters CL, Bush AI: Neurodegenerative diseases and oxidative stress. Nature Reviews Drug Discovery 2004;3(3):205-14. DOI: 10.1038/nrd1330.
https://doi.org/10.1038/nrd1330 |
| 12 | Rokad D, Ghaisas S, Harischandra DS, Jin H, Anantharam V, Kanthasamy A, Kanthasamy AG: Role of neurotoxicants and traumatic brain injury in α-synuclein protein misfolding and aggregation. Brain Res Bull 2017;133:60-70. DOI: 10.1016/j.brainresbull.2016.12.003.
https://doi.org/10.1016/j.brainresbull.2016.12.003 |
| 13 | Singh NK, Singh A, Mayank: Nuclear Factor Kappa B: A Nobel Therapeutic Target of FlavonoidsAgainst Parkinson's Disease. Comb Chem High Throughput Screen 2024;27(14):2062-77. DOI: 10.2174/0113862073295568240105025006.
https://doi.org/10.2174/0113862073295568240105025006 |
| 14 | Paterniti I, Cordaro M, Navarra M, Esposito E, Cuzzocrea S: Emerging pharmacotherapy for treatment of traumatic brain injury: targeting hypopituitarism and inflammation. Expert Opin Emerg Drugs 2015;20(4):583-96. DOI: 10.1517/14728214.2015.1058358.
https://doi.org/10.1517/14728214.2015.1058358 |
| 15 | Campolo M, Esposito E, Ahmad A, Di Paola R, Paterniti I, Cordaro M, Bruschetta G, Wallace JL, Cuzzocrea S: Hydrogen sulfide-releasing cyclooxygenase inhibitor ATB-346 enhances motor function and reduces cortical lesion volume following traumatic brain injury in mice. J Neuroinflammation 2014;11:196. DOI: 10.1186/s12974-014-0196-1.
https://doi.org/10.1186/s12974-014-0196-1 |
| 16 | Cordaro M, Impellizzeri D, Paterniti I, Bruschetta G, Siracusa R, De Stefano D, Cuzzocrea S, Esposito E: Neuroprotective Effects of Co-UltraPEALut on Secondary Inflammatory Process and Autophagy Involved in Traumatic Brain Injury. J Neurotrauma 2016;33(1):132-46. DOI: 10.1089/neu.2014.3460.
https://doi.org/10.1089/neu.2014.3460 |
| 17 | Esposito E, Cordaro M, Cuzzocrea S: Roles of fatty acid ethanolamides (FAE) in traumatic and ischemic brain injury. Pharmacol Res 2014;86:26-31. DOI: 10.1016/j.phrs.2014.05.009.
https://doi.org/10.1016/j.phrs.2014.05.009 |
| 18 | Bensalem J, Servant L, Alfos S, Gaudout D, Laye S, Pallet V, Lafenetre P: Dietary Polyphenol Supplementation Prevents Alterations of Spatial Navigation in Middle-Aged Mice. Front Behav Neurosci 2016;10:9. DOI: 10.3389/fnbeh.2016.00009.
https://doi.org/10.3389/fnbeh.2016.00009 |
| 19 | Krikorian R, Nash TA, Shidler MD, Shukitt-Hale B, Joseph JA: Concord grape juice supplementation improves memory function in older adults with mild cognitive impairment. Br J Nutr 2010;103(5):730-4. DOI: 10.1017/S0007114509992364.
https://doi.org/10.1017/S0007114509992364 |
| 20 | Dudonne S, Dal-Pan A, Dube P, Varin TV, Calon F, Desjardins Y: Potentiation of the bioavailability of blueberry phenolic compounds by co-ingested grape phenolic compounds in mice, revealed by targeted metabolomic profiling in plasma and feces. Food Funct 2016;7(8):3421-30. DOI: 10.1039/c6fo00902f.
https://doi.org/10.1039/C6FO00902F |
| 21 | Impellizzeri D, Tomasello M, Cordaro M, D'Amico R, Fusco R, Abdelhameed AS, Wenzel U, Siracusa R, Calabrese V, Cuzzocrea S, Di Paola R: MemophenolTM Prevents Amyloid-β Deposition and Attenuates Inflammation and Oxidative Stress in the Brain of an Alzheimer's Disease Rat. International Journal of Molecular Sciences 2023;24(8):6938.
https://doi.org/10.3390/ijms24086938 |
| 22 | Socci V, Tempesta D, Desideri G, De Gennaro L, Ferrara M: Enhancing Human Cognition with Cocoa Flavonoids. Front Nutr 2017;4:19. DOI: 10.3389/fnut.2017.00019.
https://doi.org/10.3389/fnut.2017.00019 |
| 23 | Brickman AM, Yeung LK, Alschuler DM, Ottaviani JI, Kuhnle GGC, Sloan RP, Luttmann-Gibson H, Copeland T, Schroeter H, Sesso HD, Manson JE, Wall M, Small SA: Dietary flavanols restore hippocampal-dependent memory in older adults with lower diet quality and lower habitual flavanol consumption. Proc Natl Acad Sci U S A 2023;120(23):e2216932120. DOI: 10.1073/pnas.2216932120.
https://doi.org/10.1073/pnas.2216932120 |
| 24 | Campolo M, Crupi R, Cordaro M, Cardali SM, Ardizzone A, Casili G, Scuderi SA, Siracusa R, Esposito E, Conti A, Cuzzocrea S: Co-Ultra PEALut Enhances Endogenous Repair Response Following Moderate Traumatic Brain Injury. Int J Mol Sci 2021;22(16). DOI: 10.3390/ijms22168717.
https://doi.org/10.3390/ijms22168717 |
| 25 | Fusco R, Gugliandolo E, Siracusa R, Scuto M, Cordaro M, D'Amico R, Evangelista M, Peli A, Peritore AF, Impellizzeri D, Crupi R, Cuzzocrea S, Di Paola R: Formyl Peptide Receptor 1 Signaling in Acute Inflammation and Neural Differentiation Induced by Traumatic Brain Injury. Biology (Basel) 2020;9(9). DOI: 10.3390/biology9090238.
https://doi.org/10.3390/biology9090238 |
| 26 | Impellizzeri D, Cordaro M, Bruschetta G, Siracusa R, Crupi R, Esposito E, Cuzzocrea S: N-Palmitoylethanolamine-Oxazoline as a New Therapeutic Strategy to Control Neuroinflammation: Neuroprotective Effects in Experimental Models of Spinal Cord and Brain Injury. J Neurotrauma 2017;34(18):2609-23. DOI: 10.1089/neu.2016.4808.
https://doi.org/10.1089/neu.2016.4808 |
| 27 | D'Amico R, Trovato Salinaro A, Fusco R, Cordaro M, Impellizzeri D, Scuto M, Ontario ML, Lo Dico G, Cuzzocrea S, Di Paola R, Siracusa R, Calabrese V: Hericium erinaceus and Coriolus versicolor Modulate Molecular and Biochemical Changes after Traumatic Brain Injury. Antioxidants (Basel) 2021;10(6). DOI: 10.3390/antiox10060898.
https://doi.org/10.3390/antiox10060898 |
| 28 | Porsolt RD, Bertin A, Blavet N, Deniel M, Jalfre M: Immobility induced by forced swimming in rats: effects of agents which modify central catecholamine and serotonin activity. Eur J Pharmacol 1979;57(2-3):201-10.
https://doi.org/10.1016/0014-2999(79)90366-2 |
| 29 | Genovese T, Siracusa R, Fusco R, D'Amico R, Impellizzeri D, Peritore AF, Crupi R, Gugliandolo E, Morabito R, Cuzzocrea S, Trovato Salinaro A, Cordaro M, Di Paola R: Atrazine Inhalation Causes Neuroinflammation, Apoptosis and Accelerating Brain Aging. Int J Mol Sci 2021;22(15). DOI: 10.3390/ijms22157938.
https://doi.org/10.3390/ijms22157938 |
| 30 | Prut L, Belzung C: The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. European Journal of Pharmacology 2003;463(1-3):3-33. DOI: 10.1016/S0014-2999(03)01272-X.
https://doi.org/10.1016/S0014-2999(03)01272-X |
| 31 | Pellow S, Chopin P, File SE, Briley M: Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. Journal of neuroscience methods 1985;14(3):149-67.
https://doi.org/10.1016/0165-0270(85)90031-7 |
| 32 | Siebold L, Krueger AC, Abdala JA, Figueroa JD, Bartnik-Olson B, Holshouser B, Wilson CG, Ashwal S: Cosyntropin Attenuates Neuroinflammation in a Mouse Model of Traumatic Brain Injury. Front Mol Neurosci 2020;13:109. DOI: 10.3389/fnmol.2020.00109.
https://doi.org/10.3389/fnmol.2020.00109 |
| 33 | Zhao P, Zhou R, Zhu XY, Liu G, Zhao YP, Ma PS, Wu W, Niu Y, Sun T, Li YX, Yu JQ, Qian ZM: Neuroprotective Effects of Lycium barbarum Polysaccharide on Focal Cerebral Ischemic Injury in Mice. Neurochem Res 2017;42(10):2798-813. DOI: 10.1007/s11064-017-2293-x.
https://doi.org/10.1007/s11064-017-2293-x |
| 34 | Araki T, Kumagai T, Tanaka K, Matsubara M, Kato H, Itoyama Y, Imai Y: Neuroprotective effect of riluzole in MPTP-treated mice. Brain Res 2001;918(1-2):176-81.
https://doi.org/10.1016/S0006-8993(01)02944-4 |
| 35 | Fleming SM, Mulligan CK, Richter F, Mortazavi F, Lemesre V, Frias C, Zhu C, Stewart A, Gozes I, Morimoto B, Chesselet MF: A pilot trial of the microtubule-interacting peptide (NAP) in mice overexpressing alpha-synuclein shows improvement in motor function and reduction of alpha-synuclein inclusions. Mol Cell Neurosci 2011;46(3):597-606. DOI: 10.1016/j.mcn.2010.12.011.
https://doi.org/10.1016/j.mcn.2010.12.011 |
| 36 | Sedelis M, Schwarting RK, Huston JP: Behavioral phenotyping of the MPTP mouse model of Parkinson's disease. Behav Brain Res 2001;125(1-2):109-25. DOI: 10.1016/s0166-4328(01)00309-6.
https://doi.org/10.1016/S0166-4328(01)00309-6 |
| 37 | Cordaro M, Siracusa R, Crupi R, Impellizzeri D, Peritore AF, D'Amico R, Gugliandolo E, Di Paola R, Cuzzocrea S: 2-Pentadecyl-2-Oxazoline Reduces Neuroinflammatory Environment in the MPTP Model of Parkinson Disease. Mol Neurobiol 2018;55(12):9251-66. DOI: 10.1007/s12035-018-1064-2.
https://doi.org/10.1007/s12035-018-1064-2 |
| 38 | Pan Z, Cui M, Dai G, Yuan T, Li Y, Ji T, Pan Y: Protective Effect of Anthocyanin on Neurovascular Unit in Cerebral Ischemia/Reperfusion Injury in Rats. Front Neurosci 2018;12:947. DOI: 10.3389/fnins.2018.00947.
https://doi.org/10.3389/fnins.2018.00947 |
| 39 | Siracusa R, Impellizzeri D, Cordaro M, Crupi R, Esposito E, Petrosino S, Cuzzocrea S: Anti-Inflammatory and Neuroprotective Effects of Co-UltraPEALut in a Mouse Model of Vascular Dementia. Front Neurol 2017;8:233. DOI: 10.3389/fneur.2017.00233.
https://doi.org/10.3389/fneur.2017.00233 |
| 40 | Srivastava P, Cronin CG, Scranton VL, Jacobson KA, Liang BT, Verma R: Neuroprotective and neuro-rehabilitative effects of acute purinergic receptor P2X4 (P2X4R) blockade after ischemic stroke. Exp Neurol 2020;329:113308. DOI: 10.1016/j.expneurol.2020.113308.
https://doi.org/10.1016/j.expneurol.2020.113308 |
| 41 | Boccella S, Iannotta M, Cristiano C, Iannotti FA, Bello FD, Guida F, Belardo C, Infantino R, Ricciardi F, Giannella M, Calignano A, Di Marzo V, Maione S, Luongo L: Treatment With 2-Pentadecyl-2-Oxazoline Restores Mild Traumatic Brain Injury-Induced Sensorial and Neuropsychiatric Dysfunctions. Front Pharmacol 2020;11:91. DOI: 10.3389/fphar.2020.00091.
https://doi.org/10.3389/fphar.2020.00091 |
| 42 | Petrosino S, Campolo M, Impellizzeri D, Paterniti I, Allara M, Gugliandolo E, D'Amico R, Siracusa R, Cordaro M, Esposito E, Di Marzo V, Cuzzocrea S: 2-Pentadecyl-2-Oxazoline, the Oxazoline of Pea, Modulates Carrageenan-Induced Acute Inflammation. Front Pharmacol 2017;8:308. DOI: 10.3389/fphar.2017.00308.
https://doi.org/10.3389/fphar.2017.00308 |
| 43 | Cordaro M, Paterniti I, Siracusa R, Impellizzeri D, Esposito E, Cuzzocrea S: KU0063794, a Dual mTORC1 and mTORC2 Inhibitor, Reduces Neural Tissue Damage and Locomotor Impairment After Spinal Cord Injury in Mice. Mol Neurobiol 2017;54(4):2415-27. DOI: 10.1007/s12035-016-9827-0.
https://doi.org/10.1007/s12035-016-9827-0 |
| 44 | Di Paola R, Impellizzeri D, Fusco R, Cordaro M, Siracusa R, Crupi R, Esposito E, Cuzzocrea S: Ultramicronized palmitoylethanolamide (PEA-um((R))) in the treatment of idiopathic pulmonary fibrosis. Pharmacol Res 2016;111:405-12. DOI: 10.1016/j.phrs.2016.07.010.
https://doi.org/10.1016/j.phrs.2016.07.010 |
| 45 | Cordaro M, Cuzzocrea S, Crupi R: An Update of Palmitoylethanolamide and Luteolin Effects in Preclinical and Clinical Studies of Neuroinflammatory Events. Antioxidants (Basel) 2020;9(3). DOI: 10.3390/antiox9030216.
https://doi.org/10.3390/antiox9030216 |
| 46 | Cordaro M, Scuto M, Siracusa R, D'Amico R, Filippo Peritore A, Gugliandolo E, Fusco R, Crupi R, Impellizzeri D, Pozzebon M, Alfonsi D, Mattei N, Marcolongo G, Evangelista M, Cuzzocrea S, Di Paola R: Effect of N-palmitoylethanolamine-oxazoline on comorbid neuropsychiatric disturbance associated with inflammatory bowel disease. FASEB J 2020;34(3):4085-106. DOI: 10.1096/fj.201901584RR.
https://doi.org/10.1096/fj.201901584RR |
| 47 | Cordaro M, Impellizzeri D, Gugliandolo E, Siracusa R, Crupi R, Esposito E, Cuzzocrea S: Adelmidrol, a Palmitoylethanolamide Analogue, as a New Pharmacological Treatment for the Management of Inflammatory Bowel Disease. Mol Pharmacol 2016;90(5):549-61. DOI: 10.1124/mol.116.105668.
https://doi.org/10.1124/mol.116.105668 |
| 48 | Impellizzeri D, Siracusa R, Cordaro M, Crupi R, Peritore AF, Gugliandolo E, D'Amico R, Petrosino S, Evangelista M, Di Paola R, Cuzzocrea S: N-Palmitoylethanolamine-oxazoline (PEA-OXA): A new therapeutic strategy to reduce neuroinflammation, oxidative stress associated to vascular dementia in an experimental model of repeated bilateral common carotid arteries occlusion. Neurobiol Dis 2019;125:77-91. DOI: 10.1016/j.nbd.2019.01.007.
https://doi.org/10.1016/j.nbd.2019.01.007 |
| 49 | Siracusa R, Paterniti I, Cordaro M, Crupi R, Bruschetta G, Campolo M, Cuzzocrea S, Esposito E: Neuroprotective Effects of Temsirolimus in Animal Models of Parkinson's Disease. Mol Neurobiol 2018;55(3):2403-19. DOI: 10.1007/s12035-017-0496-4.
https://doi.org/10.1007/s12035-017-0496-4 |
| 50 | Di Paola R, Crisafulli C, Mazzon E, Genovese T, Paterniti I, Bramanti P, Cuzzocrea S: Effect of PD98059, a selective MAPK3/MAPK1 inhibitor, on acute lung injury in mice. Int J Immunopathol Pharmacol 2009;22(4):937-50. DOI: 10.1177/039463200902200409.
https://doi.org/10.1177/039463200902200409 |
| 51 | Marklund S, Marklund G: Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem 1974;47(3):469-74. DOI: 10.1111/j.1432-1033.1974.tb03714.x.
https://doi.org/10.1111/j.1432-1033.1974.tb03714.x |
| 52 | Rajasankar S, Manivasagam T, Surendran S: Ashwagandha leaf extract: a potential agent in treating oxidative damage and physiological abnormalities seen in a mouse model of Parkinson's disease. Neurosci Lett 2009;454(1):11-5. DOI: 10.1016/j.neulet.2009.02.044.
https://doi.org/10.1016/j.neulet.2009.02.044 |
| 53 | Fan Y, Maghimaa M, Chinnathambi A, Alharbi SA, Veeraraghavan VP, Mohan SK, Hussain S, Rengarajan T: Tomentosin Reduces Behavior Deficits and Neuroinflammatory Response in MPTP-Induced Parkinson's Disease in Mice. J Environ Pathol Toxicol Oncol 2021;40(1):75-84. DOI: 10.1615/JEnvironPatholToxicolOncol.v40.i1.70.
https://doi.org/10.1615/JEnvironPatholToxicolOncol.v40.i1.70 |
| 54 | Puigventós L, Navarro M, Alechaga É, Núñez O, Saurina J, Hernández-Cassou S, Puignou L: Determination of polyphenolic profiles by liquid chromatography-electrospray-tandem mass spectrometry for the authentication of fruit extracts. Analytical and Bioanalytical Chemistry 2015;407(2):597-608. DOI: 10.1007/s00216-014-8298-2.
https://doi.org/10.1007/s00216-014-8298-2 |
| 55 | Zhang L, Wang H: Targeting the NF-E2-Related Factor 2 Pathway: a Novel Strategy for Traumatic Brain Injury. Mol Neurobiol 2018;55(2):1773-85. DOI: 10.1007/s12035-017-0456-z.
https://doi.org/10.1007/s12035-017-0456-z |
| 56 | Hanafy KA, Selim MH: Antioxidant strategies in neurocritical care. Neurotherapeutics 2012;9(1):44-55. DOI: 10.1007/s13311-011-0085-6.
https://doi.org/10.1007/s13311-011-0085-6 |
| 57 | Tebay LE, Robertson H, Durant ST, Vitale SR, Penning TM, Dinkova-Kostova AT, Hayes JD: Mechanisms of activation of the transcription factor Nrf2 by redox stressors, nutrient cues, and energy status and the pathways through which it attenuates degenerative disease. Free Radic Biol Med 2015;88(Pt B):108-46. DOI: 10.1016/j.freeradbiomed.2015.06.021.
https://doi.org/10.1016/j.freeradbiomed.2015.06.021 |
| 58 | Czabotar PE, Lessene G, Strasser A, Adams JM: Control of apoptosis by the BCL-2 protein family: implications for physiology and therapy. Nat Rev Mol Cell Biol 2014;15(1):49-63. DOI: 10.1038/nrm3722.
https://doi.org/10.1038/nrm3722 |
| 59 | Xu J, Wang H, Ding K, Zhang L, Wang C, Li T, Wei W, Lu X: Luteolin provides neuroprotection in models of traumatic brain injury via the Nrf2-ARE pathway. Free Radic Biol Med 2014;71:186-95. DOI: 10.1016/j.freeradbiomed.2014.03.009.
https://doi.org/10.1016/j.freeradbiomed.2014.03.009 |
| 60 | Li X, Wang H, Gao Y, Li L, Tang C, Wen G, Zhou Y, Zhou M, Mao L, Fan Y: Protective Effects of Quercetin on Mitochondrial Biogenesis in Experimental Traumatic Brain Injury via the Nrf2 Signaling Pathway. PLoS One 2016;11(10):e0164237. DOI: 10.1371/journal.pone.0164237.
https://doi.org/10.1371/journal.pone.0164237 |
| 61 | Nair S, Doh ST, Chan JY, Kong AN, Cai L: Regulatory potential for concerted modulation of Nrf2- and Nfkb1-mediated gene expression in inflammation and carcinogenesis. Br J Cancer 2008;99(12):2070-82. DOI: 10.1038/sj.bjc.6604703.
https://doi.org/10.1038/sj.bjc.6604703 |
| 62 | Chen F, Castranova V, Shi X, Demers LM: New insights into the role of nuclear factor-kappaB, a ubiquitous transcription factor in the initiation of diseases. Clin Chem 1999;45(1):7-17.
https://doi.org/10.1093/clinchem/45.1.7 |
| 63 | Yu M, Li H, Liu Q, Liu F, Tang L, Li C, Yuan Y, Zhan Y, Xu W, Li W, Chen H, Ge C, Wang J, Yang X: Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cell Signal 2011;23(5):883-92. DOI: 10.1016/j.cellsig.2011.01.014.
https://doi.org/10.1016/j.cellsig.2011.01.014 |
| 64 | Wu AG, Yong YY, Pan YR, Zhang L, Wu JM, Zhang Y, Tang Y, Wei J, Yu L, Law BY, Yu CL, Liu J, Lan C, Xu RX, Zhou XG, Qin DL: Targeting Nrf2-Mediated Oxidative Stress Response in Traumatic Brain Injury: Therapeutic Perspectives of Phytochemicals. Oxid Med Cell Longev 2022;2022:1015791. DOI: 10.1155/2022/1015791.
https://doi.org/10.1155/2022/1015791 |
| 65 | Verstraeten SV, Keen CL, Schmitz HH, Fraga CG, Oteiza PI: Flavan-3-ols and procyanidins protect liposomes against lipid oxidation and disruption of the bilayer structure. Free Radic Biol Med 2003;34(1):84-92. DOI: 10.1016/s0891-5849(02)01185-1.
https://doi.org/10.1016/S0891-5849(02)01185-1 |
| 66 | Verstraeten SV, Oteiza PI, Fraga CG: Membrane effects of cocoa procyanidins in liposomes and Jurkat T cells. Biol Res 2004;37(2):293-300. DOI: 10.4067/s0716-97602004000200016.
https://doi.org/10.4067/S0716-97602004000200016 |
| 67 | Oteiza PI, Erlejman AG, Verstraeten SV, Keen CL, Fraga CG: Flavonoid-membrane interactions: a protective role of flavonoids at the membrane surface? Clin Dev Immunol 2005;12(1):19-25. DOI: 10.1080/10446670410001722168.
https://doi.org/10.1080/10446670410001722168 |
| 68 | Verstraeten SV, Hammerstone JF, Keen CL, Fraga CG, Oteiza PI: Antioxidant and membrane effects of procyanidin dimers and trimers isolated from peanut and cocoa. J Agric Food Chem 2005;53(12):5041-8. DOI: 10.1021/jf058018m.
https://doi.org/10.1021/jf058018m |
| 69 | Robledinos-Antón N, Fernández-Ginés R, Manda G, Cuadrado A: Activators and Inhibitors of NRF2: A Review of Their Potential for Clinical Development. Oxid Med Cell Longev 2019;2019:9372182. DOI: 10.1155/2019/9372182.
https://doi.org/10.1155/2019/9372182 |
| 70 | Qader M, Xu J, Yang Y, Liu Y, Cao S: Natural Nrf2 Activators from Juices, Wines, Coffee, and Cocoa. Beverages 2020;6(4):68.
https://doi.org/10.3390/beverages6040068 |
| 71 | Luk KC, Kehm V, Carroll J, Zhang B, O'Brien P, Trojanowski JQ, Lee VM: Pathological alpha-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science 2012;338(6109):949-53. DOI: 10.1126/science.1227157.
https://doi.org/10.1126/science.1227157 |
| 72 | Calabrese V, Guagliano E, Sapienza M, Mancuso C, Butterfield DA, Stella AM: Redox regulation of cellular stress response in neurodegenerative disorders. Ital J Biochem 2006;55(3-4):263-82.
|
| 73 | Impellizzeri D, Campolo M, Bruschetta G, Crupi R, Cordaro M, Paterniti I, Cuzzocrea S, Esposito E: Traumatic Brain Injury Leads to Development of Parkinson's Disease Related Pathology in Mice. Front Neurosci 2016;10:458. DOI: 10.3389/fnins.2016.00458.
https://doi.org/10.3389/fnins.2016.00458 |
| 74 | Siracusa R, Scuto M, Fusco R, Trovato A, Ontario ML, Crea R, Di Paola R, Cuzzocrea S, Calabrese V: Anti-inflammatory and Anti-oxidant Activity of Hidrox((R)) in Rotenone-Induced Parkinson's Disease in Mice. Antioxidants (Basel) 2020;9(9). DOI: 10.3390/antiox9090824.
https://doi.org/10.3390/antiox9090824 |
| 75 | Logsdon AF, Lucke-Wold BP, Nguyen L, Matsumoto RR, Turner RC, Rosen CL, Huber JD: Salubrinal reduces oxidative stress, neuroinflammation and impulsive-like behavior in a rodent model of traumatic brain injury. Brain Res 2016;1643:140-51. DOI: 10.1016/j.brainres.2016.04.063.
https://doi.org/10.1016/j.brainres.2016.04.063 |
| 76 | Morabito R, Costa R, Rizzo V, Remigante A, Nofziger C, La Spada G, Marino A, Paulmichl M, Dossena S: Crude venom from nematocysts of Pelagia noctiluca (Cnidaria: Scyphozoa) elicits a sodium conductance in the plasma membrane of mammalian cells. Sci Rep 2017;7:41065. DOI: 10.1038/srep41065.
https://doi.org/10.1038/srep41065 |